Iniziamo un nuovo viaggio tra le modalità di movimento dei batteri. In questo articolo, tratteremo dei movimento flagellare. Dopo aver descritto la struttura del flagello, indicheremo i meccanismi molecolari e genetici che sono alla base della sintesi del flagello. Infine, descriveremo come il flagello permette alla cellula batterica di muoversi.
Definizione del movimento flagellellare
Il movimento dei batteri in ambienti fluidi è consentito dalla presenza sulla superficie cellulare di alcuni organelli chiamati flagelli. Per definizione, un flagello è un filamento proteico cavo e sottile che può raggiungere una lunghezza di 15 micrometri. È stato osservato che la rotazione flagellare permette ai batteri della specie Escherichia coli di nuotare ad una velocità di circa 30 micrometri al secondo; altre specie batteriche, come Bdellovibrio bacteriovorus, invece, mediante la rotazione flagellare, possono raggiungere velocità di 150 micrometri al secondo.
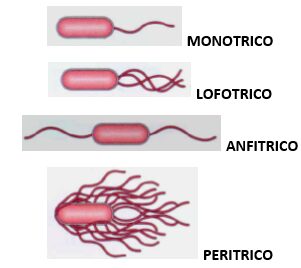
Inoltre, la quantità dei flagelli sulla superficie batterica può variare tra le diverse specie: pertanto, i batteri che possiedono un unico flagello sulla superficie cellulare sono nominati monotrichi; invece, i batteri che mostrano sulla propria superficie batterica due flagelli, ognuno distribuito ad un polo della cellula, sono detti anfitrichi; inoltre, i batteri che possiedono un ciuffo di flagelli posto ad un solo polo della superficie cellulare sono detti lofotrichi; infine, i batteri che mostrano numerosi flagelli uniformemente distribuiti sulla superficie cellulare sono nominati peritrichi (Fig. 1).
La struttura del flagello
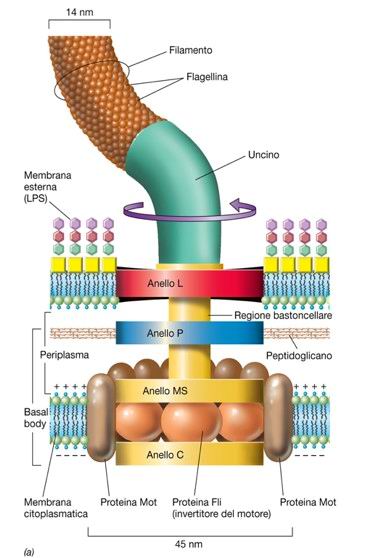
Ogni flagello è costituito da tre porzioni proteiche: il corpo basale, l’uncino e il filamento (Fig. 2). Il corpo basale ha una duplice funzione: sia permette l’ancoraggio del filamento alla cellula sia costituisce il motore del flagello. Il corpo basale flagellare è formato da numerosi anelli attraversati da una struttura cava detta bastoncello.
I batteri Gram negativi possiedono un corpo basale flagellare composto di quattro anelli, ognuno localizzato in uno strato dell’involucro cellulare: l’anello L è situato in corrispondenza della membrana esterna, l’anello P è localizzato in corrispondenza del peptidoglicano, l’anello MS è posto in corrispondenza della membrana citoplasmatica e, infine, l’anello C si estende nel citoplasma. L’anello C costituisce il rotore del movimento flagellare ed è formato da tre proteine FliG, FliM è FliN. Le proteine integrali di membrana MotA e MotB, invece, formano lo statore, la componente non ruotante del flagello.
È intuitivamente ragionevole asserire che il corpo basale del flagello dei batteri Gram positivi sia costituito solamente da due anelli: un anello è inserito nella membrana plasmatica e l’altro anello è inserito nello strato del peptidoglicano. L’uncino è una breve struttura flessibile che connette il corpo basale al filamento. Quest’ultimo è una sottile struttura tubolare costituta da circa 20000 subunità della proteina flagellina.
La sintesi dei flagelli batterici
Il processo di sintesi delle strutture flagellare è altamente regolato. Numerosi geni sia strutturali sia regolatori sono coinvolti nel processo di sintesi delle componenti del flagello.
In Escherichia coli sono circa sessanta i geni che permettono e regolano la sintesi del flagello. Tali geni sono divisi in tre classi di operoni: la prima classe di operoni ad essere trascritta è l’operone flhDC. Le proteine FlhD e FlhC formano il complesso FlhD4C2 che promuove l’espressione dei geni degli operoni di classe II. A questa classe appartengono 9 operoni i cui geni sono coinvolti nella sintesi delle proteine del corpo basale e dell’uncino. I geni di classe II, inoltre, sono gli attivatori trascrizionali per i geni di classe III. A quest’ultima classe appartengono 15 operoni i cui geni codificano sia per le proteine del filamento sia per le proteine che compongono gli anelli del motore del flagello.
La formazione del filamento del flagello avviene mediante la traslocazione delle subunità di flagellina attraverso il canale del flagello; la proteina cap, localizzata all’estremità del filamento, assicura l’assemblaggio nella corretta posizione delle subunità di flagellina.
Il movimento flagellare
Ogni batterio flagellato, sia esso monotrico, anfitrico, lofotrico oppure peritrico, può muoversi grazie alla rotazione del filamento flagellare, il quale può ruotare sia in senso orario sia in senso antiorario. Nei batteri anfitrichi, lofotrichi e peritrichi, tutti i flagelli si muovono simultaneamente nella stessa direzione. Invece nei batteri monotrichi, il flagello, se ruota in senso antiorario, permette al batterio di procedere in avanti, se, invece, ruota in senso orario, permette la retrocessione del batterio. Nei batteri lofotrichi e peritrichi, la rotazione in senso antiorario, poiché permette ai flagello di riunirsi in fasci compatti producendo una forza propulsiva, causa un movimento di avanzamento della cellula batterica (corsa), invece, la rotazione in senso orario, poiché determina l’apertura dei fasci, causa capovolgimenti casuali della cellula batterica (capriole).
Il movimento rotatorio del filamento del flagello è impartito dal motore del corpo basale. Il motore flagellare utilizza l’energia del gradiente elettrochimico della membrana cellulare per generare la rotazione del filamento. Il flusso di protoni, che attraversa le proteine MotA e MotB, causa un cambio conformazionale di MotA. MotA può muovere la proteina FliG causando la rotazione del rotore. La forza rotatoria, dunque, è trasmessa dall’anello C all’anello MS e dall’anello MS all’uncino e al filamento flagellare.
l cambiamento della rotazione del flagello dal senso antiorario al senso orario è determinato dall’interazione tra la proteina del rotore FliM e la proteina CheY del sistema di chemiotassi.
Maria Chiara Langella
Fonti
- Fonti: Biologia dei microrganismi. Dehò, Galli. Casa editrice Ambrosiana. 2016. 70 – 74.
Fonti immagini
- Immagine in evidenza: https://ibseedintorni.com
- Immagine 1: https://www.studentidibiologia.it
- Immagine 2: http://m.federica.eu