Caratteristiche
Rhizophagus irregularis (Fig. 1), noto anche come Glomus intraradices, è un fungo micorrizico arbuscolare biotrofico obbligato e forma simbiosi mutualistiche con varie specie vegetali. In particolare, questo micete è stato molto studiato per via della sua utilità nel settore agrario.
Costantemente vengono osservati effetti positivi dall’interazione di questo fungo con le piante, sia in quanto ad aumenti di resa di produzione sia come difesa da fitopatogeni (in particolare quelli del suolo). Inoltre, le continue ricerche stanno ottimizzando e rendendo più economico l’allevamento e la vendita degli inoculi.
In particolare, questa micoriza altera la produzione di essudati radicali e non solo. In questo modo la pianta riesce a difendersi da stress biotici ed abiotici.
In generale, come per altri funghi micorrizici, la creazione di questa associazione biotrofica deriva da meccanismi di segnalazione complessi e dinamici, che sono ad oggi ancora in gran parte sconosciuti. Inoltre, essi coinvolgono importanti cambiamenti di espressione genica sia dei funghi che delle piante, e tali processi sono costantemente sotto studio.
Filogenesi
| Regno: | Fungi |
| Divisione: | Glomeromycota |
| Classe: | Glomeromycetes |
| Ordine: | Glomerales |
| Famiglia: | Glomeraceae |
| Genere: | Rhizophagus |
| Specie: | R. irregularis (syn. Glomus intraradices) |
Ceppo CMCCROC7
Il ceppo di Rhizophagus irregularis CMCCROC7 è quello maggiormente utilizzato a livello commerciale. Ovviamente se viene utilizzata questa micorriza bisogna evitare l’utilizzo di fungicidi sul terreno e di fosfato solubile per circa un mese dopo la loro applicazione.
Inoltre, questo fungo non si insedia su alcune famiglie di piante: Batulaceae, Fagaceae, Pinaceae, Orchidee, Ericaceae, Crucifera, Chenopodiaceae e Caryophzllaceae.
In più, questo ceppo, è stato utilizzato per nuovi metodi di allevamento, come ad esempio utilizzando patate invece che carote. L’obiettivo di queste nuove metodologie è migliorare la produzioni di massa di spore da utilizzare come biofertilizzanti micorizzici.
Metabolismo
La simbiosi tra la pianta e R. irregularis induce l‘ospite ad accumulare prodotti vegetali di nuova formazione nel sito di infezione.
Un esempio è l’accumulo di proteine, in particolare della proteina di trasferimento dei lipidi e della fenilalanina ammoniaca liasi dopo l’insediamento della micorriza. Sempre nel pre-stadio della colonizzazione delle radici avviene l’induzione nella pianta della proteina simile all’espansina che ha il compito di segnalatore simbiontico.
Genoma
Recenti ricerche hanno anche evidenziato la capacità del fungo di indurre ampi cambiamenti del trascrittoma della pianta. Questi vengono indotti durante le fasi pre, precoce e tardiva della colonizzazione della radice.
I ricercatori hanno poi osservato l’induzione di diversi geni nella fase avanzata della colonizzazione delle radici. Questi geni sono direttamente correlati alla biosintesi della micorradicina. Questa, essendo una via dei carotenoidi, è responsabile della colorazione gialla di alcune radici micorriziche.
Inoltre, tali studi hanno dimostrato alcuni cambiamenti trascrizionali coinvolti nelle vie metaboliche che controllano gli scambi nutrizionali, il metabolismo secondario, il turnover proteico, la trasduzione del segnale, la modifica della parete cellulare, il processo di trasporto e la trascrizione.
Difesa delle piante
I cambiamenti nell’espressione genica della difesa delle piante sono essenziali per la costituzione dell’associazione tra il fungo e la pianta ospite. É da sottolineare in particolare che questi cambiamenti sembrerebbero correlati al controllo biologico conferito dai funghi micorrizici alle piante contro un’ampia gamma di agenti patogeni.
L’aumento transitorio dell’espressione dei geni di difesa delle piante è osservato specialmente nelle prime fasi della colonizzazione delle radici. Questo è poi seguito da un declino nelle fasi successive allo sviluppo.
Differenti esperimenti hanno inoltre rivelato che alcuni trascritti relativi alle risposte di difesa delle piante sono stati rilevati esclusivamente in cellule contenenti arbuscoli e, spesso, tali risposte delle piante ai funghi biotrofi obbligati sono simili a risposte delle piante ai patogeni biotrofi.
Durante questa simbiosi è stato osservato un aumento di acido jasmonico nelle radici delle piante micorriziche. Questo per via della regolazione della via dell’ossilipina, che ha un ruolo chiave nel regolare i jasmonati nella simbiosi. Infine è stato osservato lo stesso per l’acido salicilico (in particolare per due geni di carbossilmetiltranseferasi).
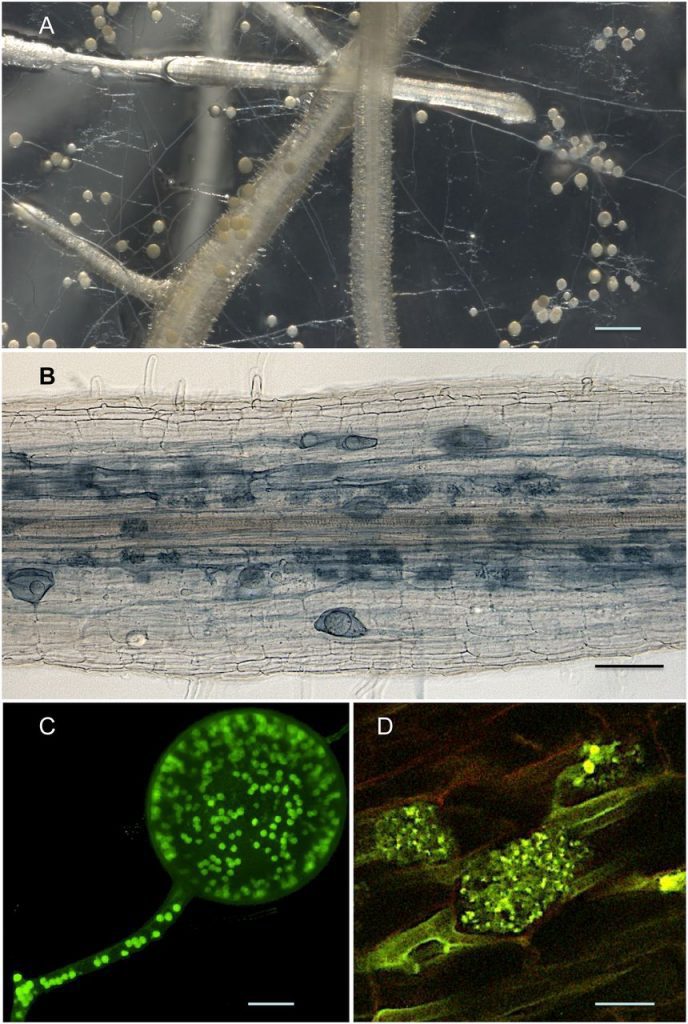
Immagini al microscopio
Nella colonizzazione fungina di R. irregularis a livello della corteccia della radice, esso produce vescicole e/o spore intraradicali e arbuscoli. La spora asessuata è multinucleata e le ife sono solitamente cenocitiche. Gli arbuscoli si presentano come strutture altamente ramificate all’interno delle cellule corticali.
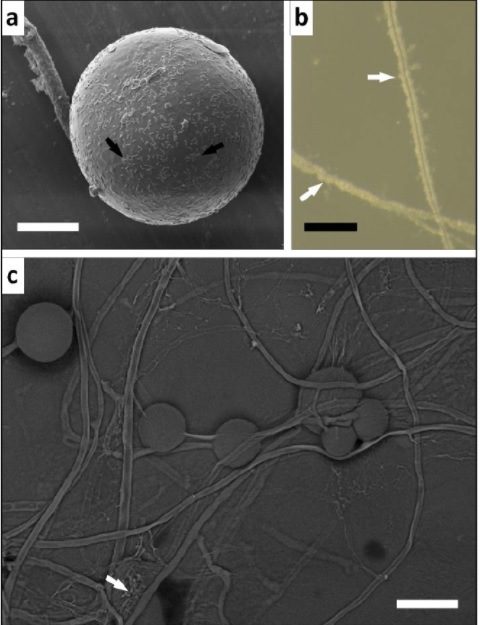
Morfologia delle colonie
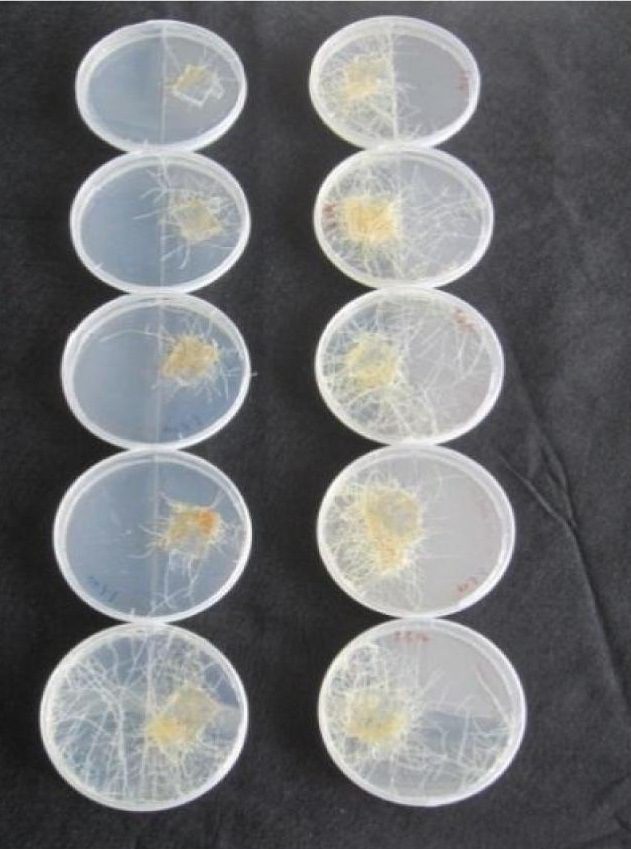
Metodi di allevamento
Rhizophagus irregularis è una delle poche specie di funghi micorrizici arbuscolari che può essere coltivata in vitro. In particolare, viene coltivato su piastre a doppio compartimento, dove il materiale fungino può essere raccolto da un compartimento di soli funghi.
Le radici delle piante ospitanti spesso “invadono” questo comparto ed è di conseguenza è necessario un regolare taglio delle radici se si vuole un materiale fungino privo di tracce di materiale vegetale. Una problematica di tale potatura è che porta all’aumento della contaminazione indesiderata da parte di altri microganismi. Per questo motivo, diversi studi si stanno concentrando per aumentare e rendere più efficiente la produzione di queste spore.
Ecologia
Rhizophagus irregularis , come gli altri funghi micorrizici arbuscolari, è un simbionte radicale obbligato. Questo significa che dipende dal proprio ospite per i nutrienti e, di conseguenza, è impossibile coltivarlo in condizioni axeniche (indipendentemente dalla pianta ospite).
Un grande vantaggio di questo fungo è inoltre la possibilità di essere utilizzato nell’idroponica grazie alla sua capacità a vivere a basse condizioni di ossigeno.
Tuttavia, negli ultimi decenni, sono stati sviluppati numerosi sistemi per la coltivazione in vitro utilizzando organi radicali o piante intere.
Ciclo vitale
La colonizzazione micorrizica arbuscolare ha inizio con un dialogo molecolare pre-simbiotico con le radici (quindi privo di contatto). Successivamente, l’ifa si differenzia con una struttura rigonfia e ramificata (ipopodio) dalla quale si svilupperà un’ifa penetrante che invaderà lo strato cellulare rizodermico.
Dopo aver attraversato gli strati cellulari esterni, il micete si diffonde in maniera longitudinale nella corteccia interna e forma gli arbuscoli (strutture altamente ramificate) all’interno delle cellule corticali. Diversi studiosi han ipotizzato che queste strutture siano il principale sito di trasferimento dei nutrienti dal fungo alla pianta.
Quindi, da un punto di vista semplificato, la colonizzazione della radice da parte di questo fungo avviene in tre fasi principali, tutte modulate da un dialogo di segnalazione tra la pianta e il micete in simbiosi.
La prima è il pre-stadio che è la fase prima del contatto del fungo con la radice, la seconda è la fase iniziale con l’avvio della colonizzazione. Infine, la terza è la fase tardiva, dove si assiste ad un’intensa colonizzazione delle radici e alla formazione di arbuscoli e vescicole.
Questa colonizzazione potrebbe differire a seconda del fatto che le piante colonizzate siano dicotiledoni o monocodtiledoni; infatti, esse differiscono sia per l’architettura dei sistemi radicali sia nel modello cellulare di alcuni tipi di radici.
Inoltre, è noto che i funghi arbuscolari modificano l’architettura delle radici dell’ospite.
Utilizzi ed azione
In generale è stato osservato che i funghi micorrizici arbuscolari possono essere utilizzati come controllo biologico dei patogeni del suolo, come Fusarium sp. (ad esempio Fusarium oxysporum f. sp. crisantemi).
In particolare, fra i patogeni del suolo controllati con l’utilizzo di tali funghi, troviamo anche i nematodi endoparassiti. Infatti, è stata studiata l’efficacia di Rhizophagus irregularis nel ridurre significativamente la presenza di Radopholus similis nelle radici di banano e di Pratylenchus coffeae nelle radici della pianta del caffè.
Nel caso dei nematodi, è stato inoltre supposto che l’azione contrastante sia dovuta ad una competizione per la penetrazione e/o i siti di nutrienti. Inoltre, anche i cambiamenti biochimici ed anatomici nella radice micorrizica, i quali sono stati osservati efficaci contro altri patogeni.
Negli ultimi anni sono state svolte diverse ricerche, per il momento solo in vitro, sulla possibile azione benefica di questi funghi su patogeni della parte aerea della pianta. E’ stata, infatti, verificata una possibile efficacia contro Phytophthora infestans.
Infatti, diversi studi hanno dimostrato che il Rhizophagus sp. è in grado di ridurre la gravità delle malattie causate da P. infestans (nota come peronospora della patata e del pomodoro), come pure da Rhizoctonia solani.
Fonti
- Glomus intraradices induces changes in root system architecture of rice independently of common symbiosis signaling (2009). Gutjahr C., Casieri L., Paszkowski U. New Phytologist, Vol. 182 (4), pp. 829-837
- Comptoir des jardins
- Use of the arbuscular mycorrhizal fungus Glomus intraradices as biological control agent of the nematode Nacobbus aberrans parasitizing tomato (2014). Marro N., Lax P., Cabello M., Doucet M. E., Becerra A. G. Agriculture, Agribusiness and Biotechnology, vol 57 (5)
- A new system using Solanum tuberosum for the co-cultivation of Glomus intraradices and its potential for mass producing spores of arbuscular mycorrhizal fungi (2013). Puri A., Adholeya A., Symbiosis Vol. 59 (2), pp. 87-97
- Giachero M. L., Marquez N., Gallou A., Luna C. M., Declerck S., Ducasse D. A. (2017) An In Vit ro Method for Studying the Three-Way Interaction between Soybean, Rhizophagus irregularis and the Soil-Borne Pathogen Fusarium virguliforme, Frontiers in Plant Science.
- Mycorrhiza induced resistance in potato plantlets challenged by Phytophthora infestans (2011) Gallou A., Mosquera L. P. H., Cranenbrouck S., Suàrez J. P., Declerck S., Physiological and Molecular Plant Pathology, Vol. 76 (1), pp. 20-26
- Effects of Rhizophagus irregularis MUCL 41833 on the reproduction of Radopholus similis in banana plantlets grown under in vitro culture conditions (2012). Koffie M. C., Vos C., Draye X., Declerck S. Mycorrhiza Vol. 23, pp. 279-288
- Impact of Rhizophagus sp. (syn. Glomus sp.) and Trichoderma harzianum on the potato resistance against Rhizoctonia solani and Phytophthora infestans, two major potato pathogens (2011). Gallou A., PhD thesis, University Catholic of Louvain.
- Use of Root Organ Cultures To Investigate the Interaction Between Glomus intraradices and Pratylenchus coffeae (2020). Elsen A., Declerck S., De Waele D., Applied and Environmental Microbiology, Vol. 69 (7)
- Transcriptional regulation of defence genes and involvement of the WRKY transcription factor in arbuscular mycorrhizal potato root colonization (2012). Gallou A., Declerck S., Cranenbrouck S. Functional & Integrative Genomics, Vol. 12, pp. 183-198
- Cost-efficient production of in vitro Rhizophagus irregularis (2017). Rosikiewicz P., Bonvin J., Sanders I. R. Vol. 27 (5), pp. 477-486
- The symbiosis with the arbuscular mycorrhizal fungus Rhizophagus irregularis drives root water transport in flooded tomato plants (2014). Calvo-Polanco M., Molina S., Zamarreno Angel Maria, Garcìa-Mina J. M., Aroca R. Plant Cell Physiol. Vol 55 (5), pp. 1017-1029
- Immagine copertina: https://commons.wikimedia.org/wiki/File:Mycorhize_dans_racine.jpg
Una piccola precisazione: Rhizophagus (syn. Rhizoglomus) irregularis non e’ sinonimo di Glomus intraradices (Rhizophagus/Rhizoglomus intraradices). Sono due specie distinte 🙂
Chiedere al Dr Fritz Oehl (che credo lavori ancora all’ Agroscope) per ulteriori dettagli.