Caratteristiche
Echinococcus multilocularis è un verme piatto (platelminta) parassita appartenente alla classe dei Cestodi, che risiede nell’intestino tenue di alcuni mammiferi carnivori, come gatti, volpi, cani e lupi. In Europa l’E. multilocularis si trova specialmente nella Francia centrale ed orientale, in Svizzera, Austria e Germania. In Asia abita nelle regioni caratterizzate dalla tundra (nel tratto che va dal mar Bianco fino allo stretto di Bering), nella parte meridionale, in particolare Turchia ed Afghanistan, e nei territori settentrionali del Giappone. Infine, è diffuso anche in varie parti dell’America, tra cui le aree subartiche dell’Alaska, il Canada nordoccidentale, fino agli stati centro-settentrionali e meridionali degli Stati Uniti.
Filogenesi
Dominio Eukaryota
Regno Animalia
Phylum Platyhelminthes
Classe Cestoda
Ordine Cyclophyllidea
Famiglia Taeniidae
Genere Echinococcus
Specie E. multilocularis
Caratteristiche strutturali
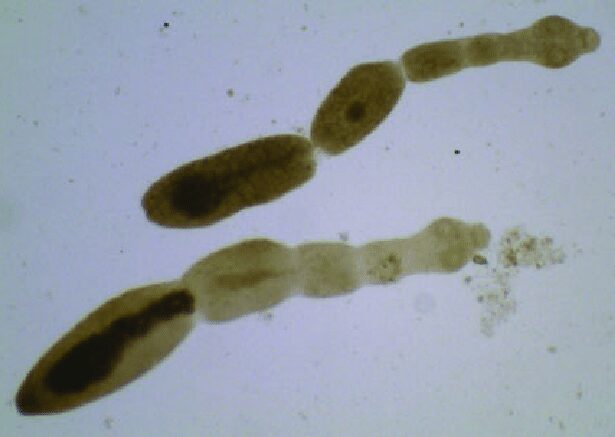
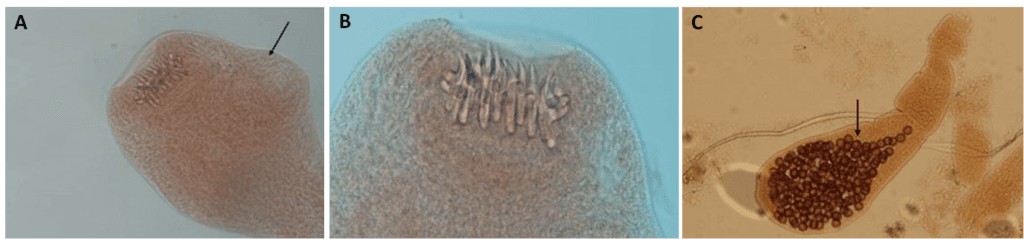
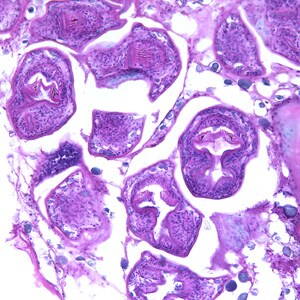
Echinococcus multilocularis è lungo circa 5-6 mm e, come gli altri cestodi, è ermafrodita (Fig.1). Ha il corpo nastriforme e segmentato, in cui si distinguono le tre componenti primarie: lo scolice, provvisto di ventose ed uncini (Fig. 2A e 2B) che permettono l’adesione all’epitelio intestinale, il collo, che ha la funzione di creare i segmenti chiamati proglottidi, e lo strobilo, una catena di proglottidi (nell’Echinococcus spp. sono solo 3, mentre nella Teania spp. possono arrivare fino a 3000), nelle quali avviene la fecondazione e la conseguente generazione delle uova (Fig. 2C). Oltre a ciò, questo platelminta manca di apparato circolatorio, respiratorio e digerente, e presenta un apparato escretore e un sistema nervoso alquanto rudimentali. Il genoma di questa classe di parassiti non contiene geni codificanti per enzimi che sintetizzano colesterolo, acidi grassi e amminoacidi, bensì i nutrienti vengono acquisiti mediante assorbimento attraverso il tegumento dotato di microvilli. Il metabolismo, anaerobio, dipende soprattutto dagli zuccheri.
Ciclo biologico e patogenesi
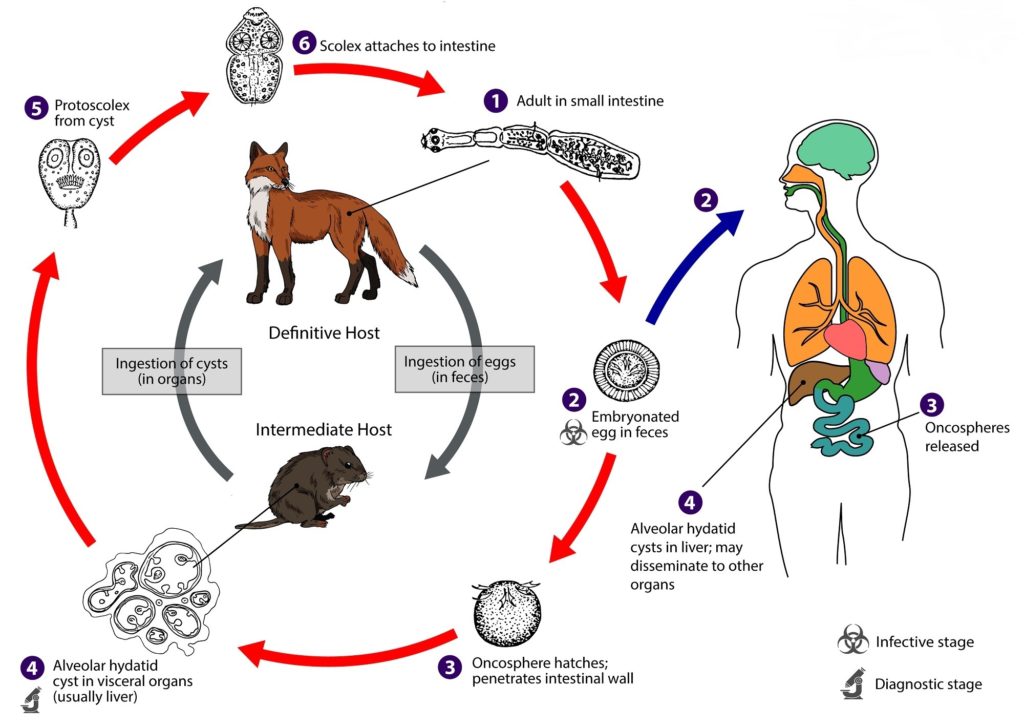
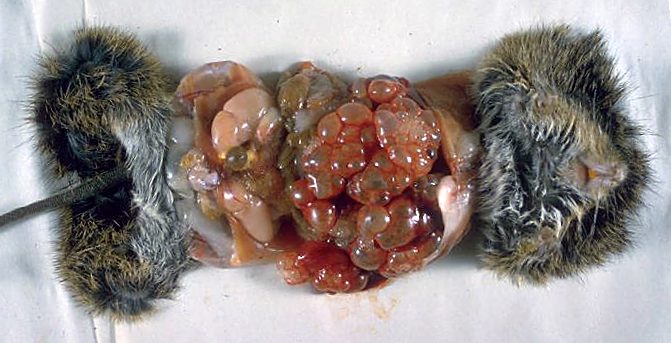
E. multilocularis è un parassita eteroxeno, ovvero sfrutta organismi appartenenti a specie diverse, e inizia il proprio ciclo vitale (Fig. 3) nel momento in cui le proglottidi, contenenti le uova mature, vengono depositate nell’ambiente mediante le deiezioni degli ospiti definitivi (volpi, cani, lupi). Queste uova racchiudono un embrione dotato di sei uncini (chiamato oncosfera o larva esacanta) e vanno a contaminare piante, funghi e frutti di bosco (fragole, mirtilli, lamponi, ribes e bacche); in seguito all’ingestione da parte di roditori selvatici, ruminanti ed esseri umani (che raffigurano gli ospiti intermedi), le uova liberano l’oncosfera che, grazie alla presenza degli uncini, riesce a perforare la parete intestinale e, attraverso i circoli ematico e linfatico, raggiunge svariati distretti dell’organismo, tra cui i polmoni, il fegato (l’organo maggiormente coinvolto), i reni, la milza, i tessuti molli, il sistema nervoso centrale, i muscoli e le ossa. A questo stadio del ciclo si verifica lo sviluppo della forma larvale (detta anche metacestode), l’idatide alveolare, che è contraddistinta da una forma “a grappolo”, in cui la ciste madre prolifera e genera le cisti figlie mediante gemmazione verso l’esterno (Fig. 4). Nelle cisti figlie si sviluppano numerosi protoscolici, che sono gli scolici prematuri. I grappoli invadono ed infiltrano gli organi parassitati provocando lesioni simil-tumorali, e si propagano come metastasi (Fig. 5).
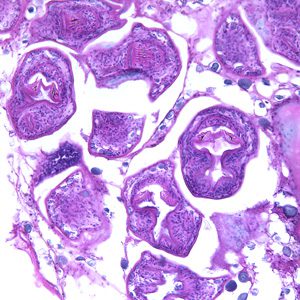
La patologia causata dall’idatide si chiama idatidosi alveolare (o echinococcosi alveolare), che rappresenta una grave zoonosi responsabile di manifestazioni tossico-allergiche, epatomegalia, dispepsie, dolore addominale, ostruzione biliare ed ascite. Se non trattata, nel 70% dei casi ha un esito tragico.
Quando i carnivori mangiano i visceri di animali infettati dalle cisti idatidee, E. multilocularis conclude il suo ciclo vitale; i protoscolici si evaginano, si attaccano alla mucosa intestinale e il verme passa alla forma adulta in un lasso di tempo che va da 32 a 80 giorni.
Rispetto all’idatide cistica (causata da E. granulosus), che è endemica in tutti i continenti, l’idatide alveolare ha una distribuzione più ristretta, confinata nell’emisfero boreale, ed è legata ad un ambiente silvestre. Si suppone che incremento del rischio di infezione per l’essere umano sia connesso all’aumento delle popolazioni di volpi e alla loro crescente urbanizzazione.
Metodi di identificazione
Indagini strumentali come la scintigrafia, la tomografia computerizzata, l’ecografia o la radiologia convenzionale possono far supporre la presenza di metacestodi nei vari tessuti e organi.
La valutazione diagnostica diretta si fonda sull’analisi morfologica ed istologica delle cisti che vengono asportate durante un intervento chirurgico. La colorazione di Gomori o quella con ematossilina-eosina mettono in evidenza la membrana cuticolare dalla ciste idatidea, composta da lamelle parallele che assumono una colorazione vivace.
Test sierologici
E’ possibile, inoltre, ottenere informazioni anche grazie ai test sierologici; è consigliato ricercare le immunoglobuline di classe E specifiche tramite il saggio REAST (Reversed Enzyme Immuno Assay) e le immunoglobuline di classe G specifiche mediante il test ELISA (Enzyme-Linked Immuno Assay). Se positive, queste ultime devono essere confermate con il Western blot. Nel caso in cui le cisti siano localizzate nel sistema nervoso centrale, le IgG si possono cercare anche nel liquido cerebrospinale. Nell’idatidosi non epatica la quantità di anticorpi è solitamente bassa.
Per la ricerca anticorpale mediante ELISA, esistono diversi antigeni, ma quelli più utilizzati sono:
- Em2, purificato da estratti tissutali del metacestode di E. multilocularis; questa molecola ha un peso molecolare di 54 kDa, e rappresenta un componente strutturale trovato esclusivamente nello strato laminato (glicocalice) dell’idatide, e non nelle oscosfere, nei protoscolici o negli stadi adulti. Il glicocalice è una struttura altamente glicosilata che si pensa giochi un ruolo importante nell’interazione ospite-parassita, proteggendo il cestode dalle reazioni immunologiche e fisiologiche dell’organismo ospite. L’antigene glicosilato Em2 (G11) raffigura il maggiore componente antigenico di Em2; studi effettuati con spettrometria di massa hanno rivelato che le componenti zuccherose sono galattosio, N-acetil-galattosamina e N-acetil-glucosamina. Inoltre, l’analisi degli amminoacidi ha segnalato una cospicua percentuale di residui di treonina e prolina, suggerendo che un nucleo con la composizione tipica delle mucine possa essere parte della struttura antigenica. Em2 (G11) induce una proliferazione non specifica dei linfociti T nei topi carenti di cellule B (μMT), e la sintesi di isotipi di IgG a bassa avidità nei topi C57BL/6. Tali proprietà sembrano indicare che Em2 (G11) è un antigene T-indipendente che potrebbe contribuire alla mancanza di protezione contro la proliferazione del metacestode. Il crescente rendimento diagnostico di Em2 ne ha permesso l’utilizzo per l’immunodiagnosi di pazienti individuali, per indagini siero-epidemiologiche su larga scala, per il monitoraggio dei pazienti dopo la chirurgia e/o la terapia e per l’identificazione di infezione da E. multilocularis negli ospiti intermedi.
- II/3 raffigura il primo gene ad essere stato clonato per scopi immunodiagnostici. Nonostante la buona performance siero-diagnostica, i problemi connessi con la sua purificazione ed il basso livello di espressione ne hanno ostacolato l’impiego nella diagnosi di routine. Allo scopo di ovviare a tale inconvenienza, è stata sub-clonata una sequenza tronca del gene originale II/3 (indicata come II/3-10), espressa come proteina ricombinante. II/3-10 è ritenuto l’epitopo diagnostico più idoneo, e ha raggiunto un successo diagnostico simile a quello di Em2.
L’impiego simultaneo di Em2 e II/3-10 nel test ELISA mostra una specificità eccellente e solo una piccola perdita nella sensibilità diagnostica. Questo saggio è attualmente disponibile in commercio per la sierodiagnosi di echinococcosi alveolare negli esseri umani (Em2plus-ELISA; Bordier Affinity Products, Crissier, Switzerland).
Terapia e profilassi
Il trattamento è basato sull’uso di farmaci antielmintici, come il mebendazolo (Vermox), l’albendazolo (Zentel) e il praziquantel (Droncit); il primo esercita la propria funzione attaccando la tubulina del parassita ed inibendo la polimerizzazione dei microtubuli, con conseguente paralisi e morte. Anche il secondo, che è un farmaco di tipo benzimidazolico, va ad impedire l’assemblaggio della tubulina, alterando la capacità di captazione ed utilizzo del glucosio. Il terzo, che è commercializzato in Italia solamente per uso veterinario, agisce aumentando la permeabilità della membrana del parassita, e ciò conduce alla paralisi.
Per la prevenzione dell’idatidosi alveolare (ma anche per quella cistica), è opportuno proibire la macellazione domiciliare senza un’ispezione veterinaria, smaltire adeguatamente i visceri del bestiame infetto nei mattatoi, al fine di evitare che i cani e i gatti li mangino. In aggiunta a ciò, la limitazione del randagismo canino ed il controllo sanitario delle greggi e dei cani esposti all’infezione rappresentano delle misure di grande importanza; bisogna rammentare infatti che le uova che vengono eliminate con le feci rimangono vitali nell’ambiente per più di due anni. Infine, ulteriori norme profilattiche sono la consumazione di funghi e frutti di bosco dopo un accurato lavaggio, e l’adozione di norme di tipo igienico nella lavorazione delle pelli.
Diversità genetica
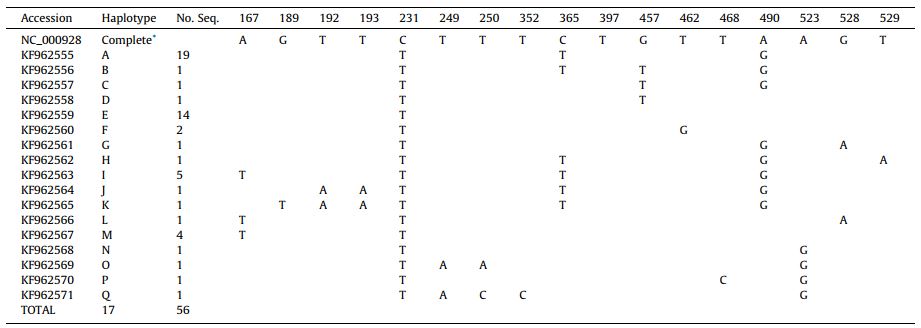
Prima di concludere mi piacerebbe citare un interessante studio del 2014 pubblicato sulla rivista International Journal for Parasitology; in questo lavoro Karen M. Gesy et al. hanno investigato la diversità genetica di E. multilocularis isolando vermi adulti da campioni fecali di 41 canidi (lupo – Canis lupus, coyote – Canis latrans, e volpe rossa – Vulpes vulpes) residenti in diverse località del Canada, tra cui Columbia Britannica, Nunavut, Alberta, Saskatchewan, Manitoba e i territori nordoccidentali. Grazie al sequenziamento di una regione di 370 nucleotidi del locus della subunità 1 della NADH deidrogenasi (nad1) mitocondriale, i ricercatori hanno identificato 17 aplotipi, che sono fondati su mutazioni a singolo nucleotide (Tabella 1).
Fonti
- Gabriella Cancrini. 2013. “Parassitologia e tecniche di laboratorio”, Lombardo Editore
- https://www.cdc.gov/parasites/echinococcosis/biology.html
- http://www.antropozoonosi.it/malattie/la-malattia-idatidea-alveolare-echinococco-multilocularis.php
- https://it.wikipedia.org/wiki/Echinococcus_multilocularis#cite_ref-1
- https://www.researchgate.net/figure/Echinococcus-multilocularis-adults-isolated-from-a-fox-in-Erzurum-province-40_fig1_305684338
- https://www.cdc.gov/dpdx/images/echinococcosis/Emultilocularis_liver_BAM1.jpg
- Zahra Heidari, Mitra Sharbatkhori, Iraj Mobedi, Seyed Hossein Mirhendi, Bahram Nikmanesh, Meysam Sharifdini, Mehdi Mohebali, Zabihollah Zarei, Kourosh Arzamani and Eshrat Beigom Kia. 2019. “Echinococcus multilocularis and Echinococcus granulosus in canines in North-Khorasan
- Province, northeastern Iran, identifed using morphology and genetic characterization of mitochondrial DNA”,Parasites Vectors
- D. Carmena, A. Benito and E. Eraso. 2007. “The immunodiagnosis of Echinococcus multilocularis infection”, European Society of Clinical Microbiology and Infectious Diseases
- Karen M. Gesy, Janna M. Schurer, Alessandro Massolo, Stefano Liccioli, Brett T. Elkin, Ray Alisauskas, Emily J. Jenkins. 2014. “Unexpected diversity of the cestode Echinococcus multilocularis in wildlife in Canada”, International Journal for Parasitology: Parasites and Wildlife 3
- https://www.my-personaltrainer.it/farmaci/vermox.html
- https://www.my-personaltrainer.it/farmaci-malattie/farmaci-schistosomiasi.html
- https://it.wikipedia.org/wiki/Albendazolo
{kind=link}