Da moltissimi anni si studiano le varie specie che abitano la terra, la loro fisionomia e il loro funzionamento, e perciò sono stati suddivisi e organizzati in gruppi sulla base delle caratteristiche comuni. Tutti gli organismi viventi sono compresi in uno dei tre grandi gruppi o domini: Bacteria, Archaea e Eukarya. Si suppone che questi tre domini derivino da un organismo antenato universale comune e che poi si sono divisi molto precocemente durante la storia della vita sulla Terra. In particolare i microrganismi batterici e archea sono procarioti unicellulari, cioè organismi in cui le cellule non hanno un nucleo delimitato da una membrana, a differenza delle cellule eucariotiche (sia unicellulari sia pluricellulari) che hanno un nucleo delimitato da una membrana. In questo articolo ci concentreremo su chi sono i batteri e come vengono classificati.
Chi sono i microrganismi batterici?
I batteri sono microrganismi unicellulari, procarioti, di dimensioni relativamente piccole, mediamente 1-2 µm e possono essere classificati in base alla morfologia (si intende la forma della cellula) in :
- cocchi, con forma sferica;
- bacilli, con forma cilindrica;
- vibrioni – spirilli: con forma ricurva o a spirale.
Le cellule di molti procarioti, in particolare i cocchi, hanno la tendenza ad associarsi in gruppi di cellule, formando strutture diverse. Ad esempio, se sono in coppia sono detti diplococchi, se formano una catenella sono chiamati streptococchi, se creano un grappolo sono detti stafilococchi.
Invece in base alla temperatura i batteri si dividono in:
- psicrofili, crescono a basse temperature (0-25°C temp. ottimale: 20- 25°C);
- mesofili, crescono a temperature intermedie (20-45°C, temp. ottimale: 30-37°C);
- termofili, crescono ad alte temperature (47-70°C, temp. ottimale: 50-55°C).
I batteri sono classificati, inoltre, in base alla loro possibilità di vivere o meno in presenza di ossigeno e vengono detti:
- batteri aerobi obbligati, crescono solo in presenza di O2;
- batteri aerobi facoltativi, possono vivere anche in assenza di ossigeno, ma la loro crescita è più veloce in presenza di ossigeno;
- batteri anaerobi obbligati, vivono e crescono solo in assenza di ossigeno.
Un ulteriore classificazione viene fatta in base al loro metabolismo e comprende:
- autotrofi, sintetizzano composti organici di cui necessitano a partire da composti inorganici semplici (CO2 e HO2);
- eterotrofi, si nutrono di composti organici.
Qual è la loro struttura ?
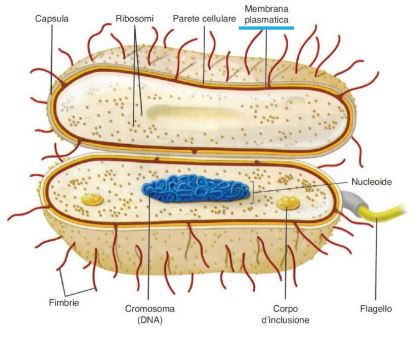
Le cellule batteriche sono formate da strutture fondamentali, presenti in tutti i tipi di batteri, e da strutture facoltative, presenti solo in determinate specie batteriche. Dall’interno verso l’esterno è formata da nucleo, citoplasma, membrana plasmatica, parete cellulare che rappresentano le strutture fondamentali, mentre le strutture accessorie sono la capsula, membrana esterna, i flagelli e plasmidi.
Il citoplasma è un gel colloidale che contiene 80% di acqua nel quale sono disciolte molecole di varia natura come proteine, zuccheri, lipidi e sono presenti anche i ribosomi ovvero organelli deputati alla sintesi proteica. Nel citoplasma troviamo il genoma che è costituito da un unico filamento circolare non racchiuso da una membrana nucleare ma che risiede in una zona detta nucleoide. Di solito questo DNA è in stretta associazione con particolari regioni della membrana plasmatica (mesosomi), dove risiedono gli enzimi per la replicazione batterica e per la produzione di energia.
Membrana citoplasmatica
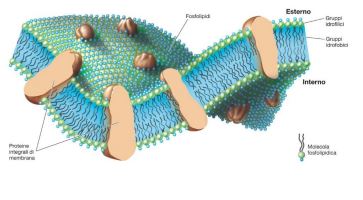
Come tutte le cellule anche i microrganismi batterici sono delimitati da una membrana citoplasmatica che separa la cellula dall’ambiente esterno. Si presenta come un doppio strato fosfolipidico in cui sono immerse proteine e glicolipidi. I fosfolipidi in soluzione acquosa si aggregano e tendono a formare spontaneamente la struttura a doppio strato, questo perché i fosfolipidi sono formati da una componente idrofobica (gli acidi grassi) che è rivolta all’interno e da una componete idrofilica (glicerofosfato) che è rivolta all’esterno o verso il citoplasma. A differenza delle cellule eucariotiche, la membrana non presenta steroli ma molecole chiamate opanoidi, che rafforzano la membrana rendendola più rigida. La membrana svolge molti ruoli, primo fra tutti quello di ”portinaio”, cioè controlla il passaggio di sostanze dal suo interno verso l’esterno e viceversa.
Come fanno le sostanze a passare attraverso la membrana? E soprattutto come fa la membrana a decidere quali sostanze devono passare e quali ad essere bloccate?
Le cellule, per poter svolgere le proprie attività, devono compiere continuamente due azioni, cioè di rifornirsi di sostanze nutritive dall’esterno e eliminare le sostanze di scarto mantenendo l’equilibrio cellulare. Gli scambi di molecole tra le cellule e l’ambiente esterno si verificano appunto attraverso la membrana perché riveste tutta la cellula. Infatti il passaggio di sostanze specifiche è consentito da specifici canali proteici, mentre altre possono attraversare la membrana indisturbate. Pertanto, è possibile distinguere due tipi di trasporto delle sostanze: trasporto attivo e trasporto passivo.
Sistemi di trasporto passivo
Il trasporto passivo avviene senza consumo di energia e permette il passaggio di sostanze come l’ossigeno, l’acqua o molecole idrofobiche secondo gradiente di concentrazione (cioè dal mezzo dove la concentrazione è maggiore al mezzo dove è minore). Ci sono due tipi di trasporto passivo: la diffusione semplice e la diffusione facilitata. Nella diffusione semplice possono attraversare liberamente la membrana solo solo piccole molecole idrofobe come O2, N2 ,H2O mentre nella diffusione facilitata è mediata da canali proteici che facilitano e velocizzano il passaggio di specifiche molecole o ioni.
Sistemi di trasporto attivo
Invece il trasporto attivo avviene mediante il consumo di energia sotto forma di ATP e contro gradiente di concentrazione e perciò si avvale di proteine specifiche. Nei batteri esistono almeno tre classi di sistemi di trasporto: il trasporto semplice, la traslocazione di gruppo e il sistema ABC.
Il trasporto semplice avviene grazie a delle proteine dette trasportatori che possono essere di tre tipi ovvero uniporter se viene trasporta una sola specie molecolare; symporter se trasporta contemporaneamente due molecole nella stessa direzione e antiporter se trasporta due molecole ma in direzioni opposte.
La traslocazione di gruppo è un sistema di trasporto in cui la sostanza trasportata viene modificata chimicamente durante l’attraversamento della membrana. Uno dei sistemi più studiati riguarda il trasporto degli zuccheri (glucosio, mannosio e fruttosio) che vengono fosforilati durante il trasporto grazie al sistema delle fosfotransferasi.
Il sistema di trasporto ABC (ATP-Binding Cassette) trasportano composti organici come zuccheri e amminoacidi o anche composti inorganici come solfato e fosfato. La caratteristica principale è la presenza di tre gruppi di proteine : una proteina periplasmatica che lega il substrato, una proteina transmembrana che forma il canale di trasporto e infine una proteina citoplasmatica che idrolizza l’ATP fornendo l’energia necessaria per l’evento di trasporto.
La parete cellulare dei microrganismi batteri
All’esterno della membrana ritroviamo una struttura caratteristica ovvero la parete batterica che oltre a proteggere la cellula dalla lisi osmotica, le conferisce forma e rigidità evitando che si danneggi quando si trova in un ambiente con ridotta pressione osmotica. E’ molto importante sottolineare, a tal proposito, che i batteri possono presentare due differenti strutture di parete cellulare. Questa distinzione è basata sulla loro reazione alla colorazione di Gram dovuta proprio alla diversa composizione chimica della parete e perciò i batteri si possono distinguere in due grandi gruppi:
- batteri Gram-positivi in cui la parete è molto spessa, formata da numerosi strati di peptidoglicano;
- batteri Gram-negativi in cui la parete risulta più complessa e sottile, circondata però da una membrana esterna.
La componente fondamentale della parete è un polimero macromolecolare chiamato peptidoglicano, ovvero uno strato rigido che è responsabile della resistenza della parete. Esso è composto da:
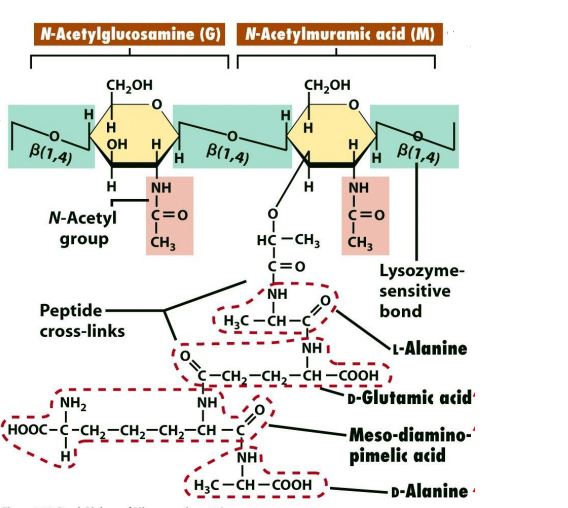
- una parte glicanica (costituita da una alternanza di N-acetilglucosamina ed acido N-acetilmuramico).
I due zuccheri sono legati mediante un legame glicosidico β1,4, che può diventare bersaglio per il lisozima. Questo enzima taglia i legami β1,4-glicosidici tra i due zuccheri, indebolendo così la parete. Al gruppo carbossilico dell’acido muramico è legato un tetrapeptide i cui amminoacidi sono rappresentati ,nell’ordine, da L-alanina, acido D-glutamico, L-lisina o acido mesodiaminopimelico e D-alanina. - una parte peptidica che forma legami crociati in grado di unire polimeri adiacenti di peptidoglicano. Infatti i polimeri sono collegati tra loro in corrispondenza delle catene amminoacidiche mediante legami che si stabiliscono tra D-alanina terminale di un tetrapeptide e l’aminogruppo della lisina mediante un ponte pentaglicinico se si tratta di batteri Gram+ o tra la D-alanina e l’acido mesodiaminopimelico se si tratta di batteri Gram-.
La parete dei batteri Gram positivi
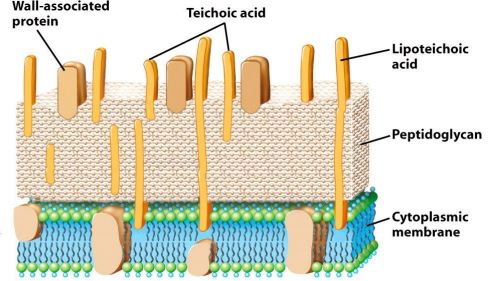
Nei batteri Gram positivi la parete è costituita da circa il 90% di peptidoglicano e perciò appare notevolmente più spessa. In essa sono presenti acidi teicoici e acidi lipoteicoici, che sono polimeri di glicerolfosfato o ribitolfosfato e sono legati covalentemente al peptidoglicano. Dato che i fosfati sono carichi negativamente, gli acidi teicoici sono responsabili della carica elettrica negativa complessiva della superficie cellulare. Gli acidi teicoici svolgono anche la funzione di legare gli ioni Ca2+ e Mg2+ per il trasporto all’interno della cellula e sono altamente antigenici perché rappresentano la porzione più esterna della cellula. Alcuni acidi teicoici sono legati covalentemente ai lipidi di membrana e perciò vengono definiti acidi lipoteicoici che servono ad ancorare la parete cellulare alla membrana citoplasmatica sottostante.
La parete dei batteri Gram negativi
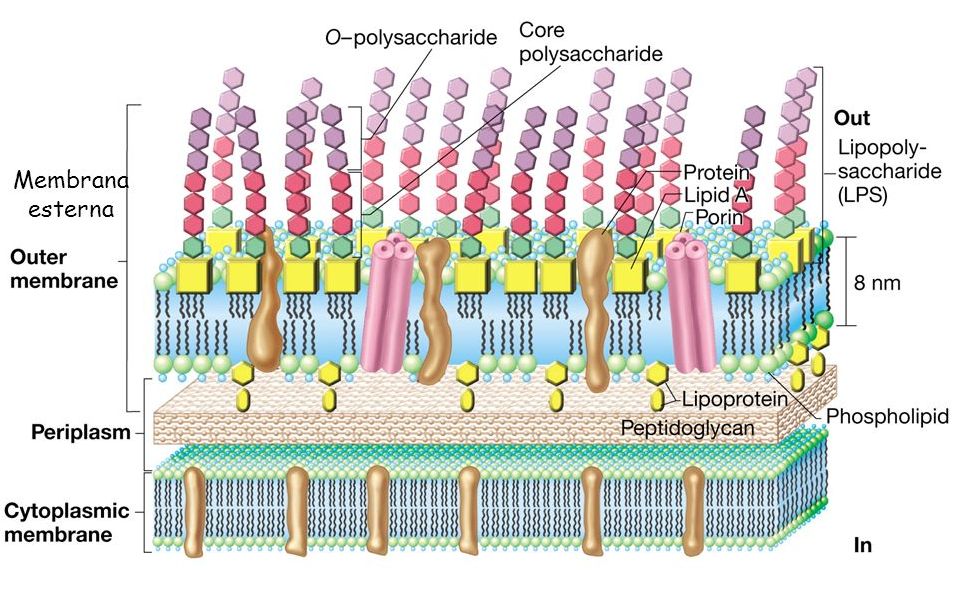
Nei batteri Gram negativi il peptidoglicano costituisce solo il 10% della totalità della parete. La maggior parte di essa è invece costituita dalla membrana esterna. Essa ha una struttura bilaminare e asimmetrica poiché è composta da un monostrato fosfolipidico più interno e uno strato più esterno di lipopolisaccaride (LPS). Quest’ultimo consiste di tre porzioni che dall’interno all’esterno sono:
- una porzione lipidica, denominata lipide A, che rappresenta l’endotossina vera e propria. E’ un glicolipide costituito da un disaccaride di glucosamina fosforilata avente legati degli acidi grassi attraverso legami esterei. Il disaccaride è legato al core polisaccaridico attraverso chetodeossiottonato (KDO);
- una corta catena di zuccheri, che forma la parte centrale della molecola, caratterizzata dalla presenza di due zuccheri che sono l’acido chetodeossiottonato e vari zuccheri a sette atomi di carbonio;
- una lunga catena polisaccaridica, denominato polisaccaride O. Esso è costituito dalla ripetizione di una serie di zuccheri in modo da formare un lungo polisaccaride O. E’ la parte più variabile e inoltre essendo la parte più esterna, funge da Antigene O che verrà riconosciuto dagli anticorpi.
La membrana esterna dei batteri è relativamente permeabile a piccole molecole idrofile grazie alla presenza di molte proteine transmembrana in particolare le porine, che formano canali permettendo il passaggio di molecole attraverso la membrana.
Strutture cellulari di superficie dei microrganismi batterici
Le cellule possono avere ulteriori componenti definite accessorie poiché non sono indispensabili per la sopravvivenza della cellula ma conferiscono solamente una caratteristica aggiuntiva. Tra le varie componenti ci sono i flagelli, pili (o fimbrie) e capsula.
I microrganismi batterici possiedono la capacità di muoversi attivamente per mezzo di particolari estroflessioni cellulari definiti flagelli. A seconda del numero e della posizione dei flagelli, si dividono in: monotrichi (quando possiedono un unico flagello), anfitrichi (quando sono dotati di due flagelli disposti ai due poli della cellula), lofotrichi (quando i flagelli sono numerosi e tutti localizzati all’estremità tanto da formare un ciuffo), peritrichi (quando tutta la superficie esterna è ricoperta da flagelli).
I flagelli hanno una struttura filamentosa e sono costituiti da subunità proteiche elicoidali contenenti flagellina. Inoltre in ciascun flagello si possono riconoscere tre parti:
- il filamento, che è la porzione sporgente;
- un gancio che unisce il filamento alla superficie cellulare;
- corpo basale, che ancora il flagello alla parete e alla membrana citoplasmatica.
All’interno del corpo basale viene generata l’energia necessaria a far muovere il flagello in senso orario o antiorario. L’andamento dei movimenti è influenzato dagli stimoli captati dai recettori posti sulla superficie del batterio; se questi avvertono la presenza di nutrienti si genera un movimento propulsivo attivo; viceversa, se il segnale captato è nocivo, si ha chemotassi negativa e il batterio si allontana.
Fimbrie e capsula
Le fimbrie e i pili sono strutture filamentose disposte sulla superficie della cellula che consentono alla cellula di aderire a superfici di cellule ospiti. A differenza delle fimbre, i pili sono più lunghi e sottili e intervengono nei processi di scambio genetico (coniugazione) fra i microrganismi batterici.
In base alla loro composizione chimica i rivestimenti esterni alla cellula batterica sono definiti capsula se lo strato è organizzato in una fitta matrice che non permette il passaggio di piccole molecole o strato mucoso se lo strato è più facilmente deformabile e pertanto non è in grado di impedire il passaggio di particelle. La capsula può svolgere varie funzioni ovvero favorisce l’adesione del batterio a determinate superfici o ad altri batteri, ma costituisce anche un fattore di virulenza in quanto protegge la cellula dalla fagocitosi.
Plasmidi: trasferimento di informazione genetica tra microrganismi batterici
I plasmidi sono piccoli elementi genetici in grado di replicare autonomamente dal cromosoma. Essi includono delle regioni che codificano per geni non essenziali alla sopravvivenza. A differenza del DNA cromosomale che è essenziale per la crescita batterica, quello plasmidico conferisce particolari caratteristiche come la resistenza ad un antibiotico, attivazione di vie metaboliche o produzione di tossine. I plasmidi vengono classificati in diverse tipologie in base al contenuto genico e quindi alla funzione in: plasmidi metabolici (contengono geni che conferiscono particolari capacità metaboliche alla cellula), plasmidi F (contengono geni che codificano per le proteine dei pili sessuali e quindi sono coinvolti nel processo di trasferimento genetico), plasmidi R (contengono geni codificanti proteine che demoliscono o modificano gli antibiotici), plasmidi di virulenza (contengono geni che conferiscono caratteristiche virulenti al ceppo come ad esempio la capacità di produrre tossine).
Grazie alle piccole dimensioni vengono utilizzati come vettori di clonaggio cioè molecole che permettono il trasferimento di sequenze di DNA da una cellula all’altra. Questi vettori si ottengono ingegnerizzando plasmidi naturali al fine di conferire loro particolari requisiti quali un’origine di replicazione che gli permetta di replicarsi indipendentemente dal cromosoma, devono contenere un marker selettivo per distinguere le cellule in ossesso del plasmide da quelle che se sono prive e infine devono contenere una regione chiamata polylinker nella quale può essere inserito il DNA estraneo.
Conclusione
Quindi avere una struttura semplice non è sinonimo di semplicità anzi, la capacità di essere molto versatili e potersi adattare al meglio ai cambiamenti, li rende estremamente complessi permettendo di confrontarci ogni giorno con nuove conoscenze.
Fonti
- www.chimica-online.it/biologia/batteri.htm
- www.istitutoagrariosartor.edu.it/wp-content/uploads/2013/11/Batteri.pdf
- www.dbcf.unisi.it/sites/st13/files/allegatiparagrafo/27-06-2013/i-batteriologia_generale_2013.pdf
- pubmed.ncbi.nlm.nih.gov/24024634/
- www.sciencedirect.com/science/article/abs/pii/S0723202017300747
- scuolamedicina.unich.it/sites/sc01/files/1_inferm_ass_cellulabatterica.pdf
Fonti immagini
- Figura 1: scuolamedicina.unich.it
- Figura 2: scuolamedicina.unich.it
- Figura 3:www.dbcf.unisi.it
- Figura 4: www.dbcf.unisi.it
- Figura 5:www.slideserve.com