I bacini anossici ipersalini (Deep Hypersaline Anoxic Basins – DHABs) sono ambienti estremi situati in diversi punti profondi dei mari e degli oceani, caratterizzati da condizioni di elevata salinità, ossigeno assente ed alte pressioni. Solo pochi microrganismi poliestremofili riescono a sopravvivere in questi ambienti così ostili alla vita come la conosciamo.
Struttura di un bacino anossico ipersalino
Un DHABs (detto anche salina o brina) è una particolare formazione sottomarina, estrema sotto ogni punto di vista. Salinità molto superiore all’acqua di mare al di sopra di esso, ossigeno praticamente nullo e pressioni di centinaia di volte superiori a quella atmosferica: sono queste le principali forzanti ambientali presenti in questi ecosistemi peculiari; sono inoltre spesso presenti alte concentrazioni di metano ed acido solfidrico. Tutte queste condizioni li rendono probabilmente l’habitat più estremo presente sul nostro pianeta.
Se scendiamo lentamente annotando la salinità (S) metro dopo metro, ad esempio impiegando una sonda CTD, otteniamo il profilo verticale di salinità della colonna d’acqua. Nelle acque profonde la salinità è un parametro costante, e come tale non ci accorgeremmo di essere in presenza di un DHAB fino a quanto il valore di S non schizzerà verso valori alti: abbiamo superato l’aloclino. Esso indica una repentina variazione di salinità all’interfaccia tra le acque “normali” e quelle ipersalate. A causa della diversa densità delle due masse d’acqua, esse non si mischiano. Nonostante le acque profonde tendano ad essere ricche di ossigeno, le acque di un DHAB ne sono completamente prive, isolate dall’aloclino.

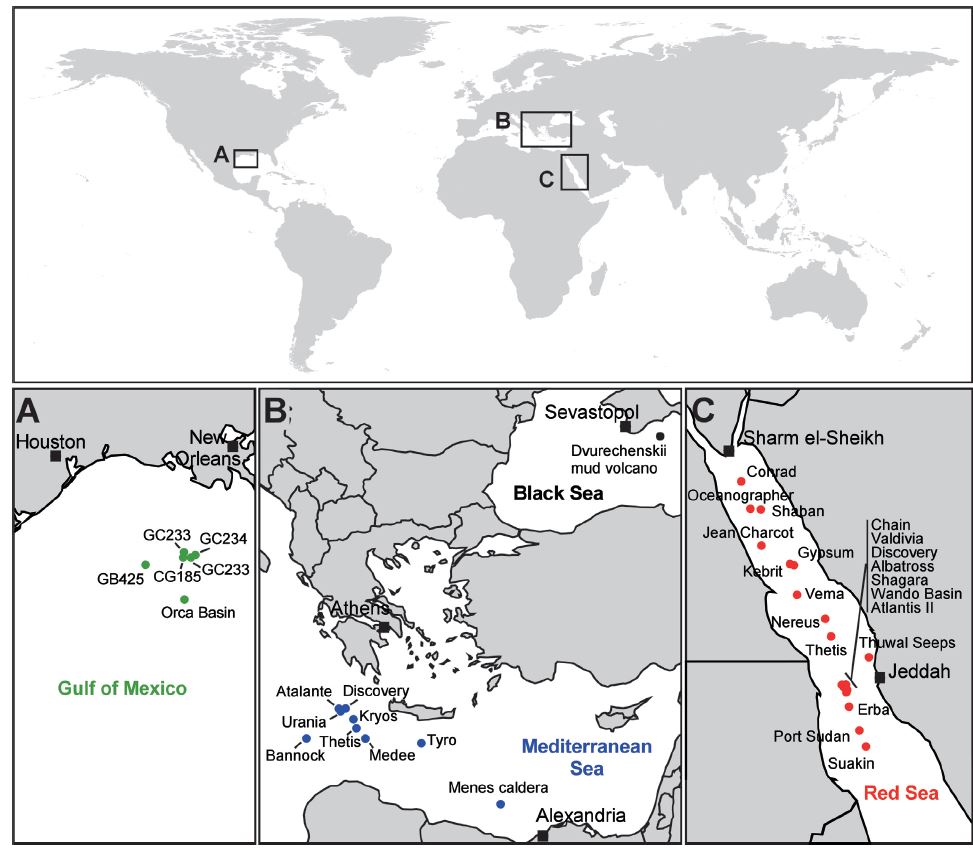
Localizzazione dei bacini anossici ipersalini
Affinché si venga a creare un ambiente estremo come un DHAB, sono necessari tutta una serie di eventi geologici in sequenza: innanzitutto si deve creare un enorme deposito evaporitico di sale, spesso originato dalla completa (o quasi) evaporazione di un tratto di mare; successivamente, si deve creare il bacino tramite processi tettonici e tale bacino deve entrare in contatto con il deposito di sale.
Oltre 5 milioni di anni fa, sulla fine del Miocene, lo stretto di Gibilterra si chiuse: il Mediterraneo, da sempre un bacino con bilancio negativo, iniziò a evaporare, creando un unico ed immenso deserto di sale, un evento chiamato “Crisi di salinità del Messiniano“. Ciò perdurò fino a che le acque atlantiche riuscirono a penetrare in Mediterraneo con la cosiddetta “Alluvione Zancleana“, dove in pochi anni le acque ricoprirono il deserto creatosi in varie migliaia.
Alla luce di ciò, non stupisce che il Mediterraneo ospiti vari DHABs, in particolare nel più profondo bacino orientale; il Golfo del Messico e il Mar Rosso sono gli altri principali bacini dove ritroviamo questi ambienti estremi. Questi tre mari sono accomunati infatti dall’essere bacini più o meno chiusi, condizione basilare per la formazione dei depositi evaporitici di sale.
La vita nei bacini anossici ipersalini
Un habitat così estremo è senza dubbio agli antipodi di ciò che possiamo associare alla vita; ma anche qui, essa ha trovato una via. Batteri ed Archea alofili ed anaerobi sono stati ritrovati all’interno dei DHAB, e sulle sue “rive” sono stati ritrovati mat batterici estesi; macrofauna di vario tipo, da policheti ad artropodi, vivono associati a queste mat microbiche, nutrendosi di biofilm e della “neve marina”. Inoltre, un particolare loricifero, Spinoloricus cinziae, è stato identificato all’interno del DHAB “L’Atalante” del Mediterraneo; si tratta del primo metazoo scoperto che non richiede ossigeno in nessuno stadio del ciclo vitale.

La produttività risulta essere più elevata a livello dell‘interfaccia tra le acque ipersaline e quelle marine, dove si ha l’accumulo del materiale disciolto e particolato che discende dalla superficie e non riesce ad oltrepassare l’aloclino. Assistiamo ad una vera e propria stratificazione batterica lungo l’interfaccia, che può avere anche un altezza di metri; l’eterogeneità delle condizioni infatti funziona da hotspot per la biodiversità batterica all’interno dell’interfaccia stessa.
Buona parte della biodiversità batterica sembra però anche essere associata a simbiosi di vario genere in vicinanza del bordo del DHABs. Questi eventi simbiotici e la diversità virale sono ancora grosse incognite da sbrogliare per avere un’estesa comprensione di questi ecosistemi estremi.

Una biochimica estrema e poco compresa
Si era inizialmente ipotizzato che l’intera produttività batterica potesse basarsi sulla decomposizione della materia in sprofondamento, salvo poi aprire all’ipotesi della chemiosintesi; come detto precedentemente, l’aloclino impedisce il passaggio di buona parte della materia organica. Tra le vie chemiosintetiche proposte si ha la metanogenesi, ipotesi legata all’osservazione di metano all’interno dei DHABs; in particolare il metano sarebbe generato a partire dalle metilammine, prodotte dalla fermentazione di molecole impiegate per la protezione dall’elevata salinità (glicina-betaina). Tale ipotesi è ancora da provare, dato che nessun batterio in grado di eseguire tale pathway è stato ancora isolato da alcun DHABs.
Per quanto riguarda l’azoto, sono state ritrovate tracce di denitrificazione a livello dell’interfaccia tra acqua ipersalina e marina. La denitrificazione è un importante sorgente di azoto nei DHABs, in grado di liberare fino all’82% dell’azoto molecolare presente. Un fatto curioso è che, mentre buona parte dell’ammoniaca è presente ad elevate concentrazioni nella parte superiore dell’interfaccia, le nitrogenasi erano molto più rappresentate nei livelli più profondi, si pensa impiegate nei pathway per la biosintesi di molecole osmo-protettrici.
L’attuale impossibilità di coltivare organismi provenienti da questi ambienti e le difficoltà nel reperire DNA ambientale intatto sono due grossi freni allo studio di questi microrganismi così estremi ed affascinanti. Inoltre, ogni bacino presenta caratteristiche peculiari, difficilmente generalizzabili ad altri ambienti dello stesso tipo.

Fonti
- Danovaro G. et al., 2010 – The first metazoa living in permanently anoxic conditions
- Yakimov M. M. et al., 2013 – Microbial life in the Lake Medee, thelargest deep-sea salt-saturated formation
- Gambi C., Danovaro R., Kristensen R. M., 2014 – Spinoloricus cinziae (Phylum Loricifera), a new species from a hypersaline anoxic deep basin in the Mediterranean Sea
- Merlino G. et al., 2018 – Microbial ecology of deep-sea hypersaline anoxic basins