Caratteristiche
Steinernema carpocapsae (Fig. 1) è un verme appartenente al phylum dei nematodi (vermi cilindrici, in contrapposizione ai vermi piatti) ed è un parassita entomopatogeno, ovvero ha come tropismo gli insetti, per i quali risulta letale. Lo S. carpocapasae vive in associazione mutualistica con Xenorhabdus nematophila (Fig. 2), un batterio Gram-negativo di forma bastoncellare, residente nell’intestino del nematode. Nel momento in cui infettano un insetto, i due microorganismi prevalgono sull’immunità dell’ospite e lo uccidono rapidamente; il batterio prolifera e trasforma i tessuti della vittima in una fonte di nutrimento per il verme, il quale va incontro a crescita e riproduzione. Grazie a questa singolare caratteristica, S. carpocapasae viene impiegato come agente di controllo biologico nell’agricoltura e nell’eliminazione degli insetti dannosi per le colture come lepidotteri, coleotteri o ditteri (di questa applicazione parleremo in dettaglio più avanti).


S. carpocapasae è in grado di percepire la produzione di anidride carbonica e utilizzare gli spiracoli (aperture per l’apparato respiratorio degli insetti) come chiave di entrata nell’organismo ospite; è maggiormente efficace a temperature che vanno da 22° C e 28° C.
Questo nematode, come anche gli altri della famiglia Steinernematidae, è ubiquitario e risiede esclusivamente nel suolo. Inoltre, è stato isolato da ogni continente abitato e da una vasta gamma di terreni ecologicamente dissimili, tra cui i campi coltivati, le foreste, i deserti, le praterie e persino le spiagge oceaniche.
Filogenesi
Dominio Eukaryota
Regno Animalia
Phylum Nematoda
Classe Secernentea
Ordine Rhabditida
Famiglia Steinernematidae
Genere Steinernema
Specie S. carpocapasae
Morfologia e ciclo vitale
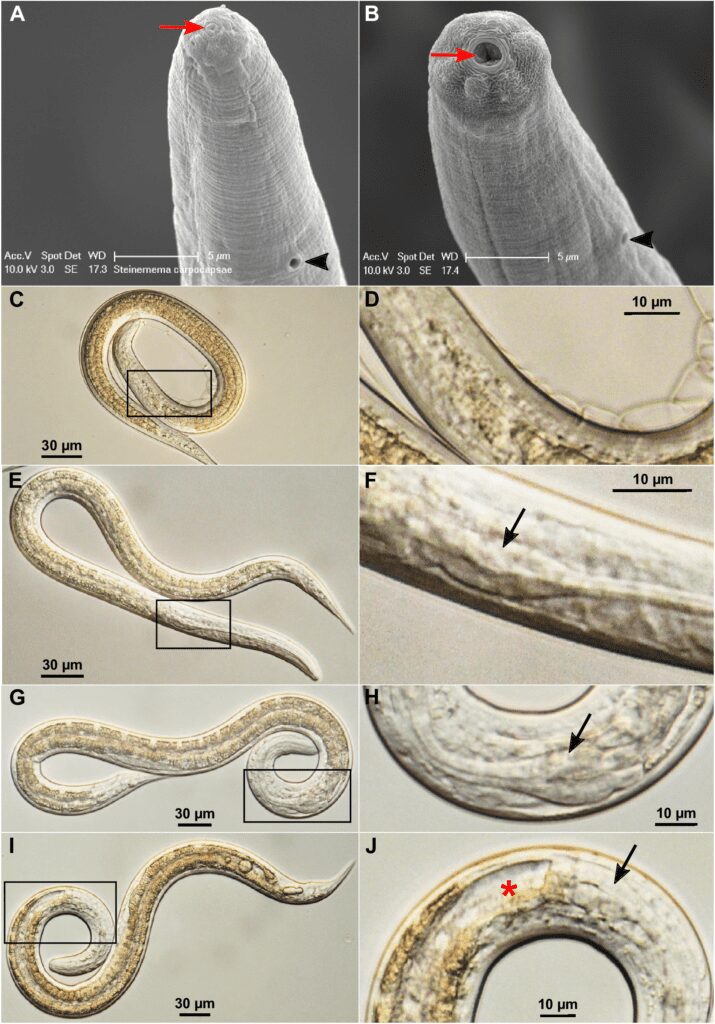
Ricordiamo che nel ciclo di sviluppo nei nematodi il raggiungimento dello stadio adulto richiede normalmente il passaggio attraverso quattro fasi larvali (L1, L2, L3 e L4); in alcuni nematodi che si comportano come parassiti intestinali degli esseri umani (come Ancylostoma duodenale, Necator americanus, Enterobius vermicularis e Strongyloides stercoralis), la larva L3 raffigura quella infettante. Lo stesso discorso riguarda lo S. carpocapsae; quando raggiunge lo stadio di infettivo immaturo (infective juvenile, IJ), che non è altro che un terzo stadio larvale modificato, riesce a vivere libero nell’ambiente esterno, è bloccato dal punto di vista dello sviluppo, non si nutre e cerca un insetto da parassitare. Durante questa fase, il nematode misura tra 0,4 e 1,5 mm di lunghezza, ha la bocca sigillata e la faringe ridotta.
Non appena entra in un organismo ospite, nella larva infettante ricomincia il ciclo di maturazione; la transizione da larva infettiva libera a parassita attivo si chiama in svariati modi, tra cui riattivazione, recupero dell’alimentazione, ripresa dello sviluppo o avvio dello sviluppo, ristabilimento e attivazione. Questo processo di mutamento comporta due importanti cambiamenti fisiologici e morfologici:
- L’apertura della cavità orale, che non è più sigillata (Fig. 3A e 3B);
- L’espansione del bulbo faringeo, che diventa facilmente osservabile al microscopio (Fig. 3F, 3H e 3J). Allo stadio IJ è quasi invisibile (Fig. 3D).
La larva allo stadio infettivo immaturo penetra nell’insetto ospite mediante varie aperture, cioè gli spiracoli, la bocca, l’ano o attraverso le membrane intersegmentali della cuticola, per poi raggiungere l’emocele (o emoceloma), rappresentante il sistema circolatorio. In seguito all’entrata nell’insetto, si verificano una serie di eventi: il parassita riprende il suo sviluppo, compie la muta e inizia a ingerire l’emolinfa. Ciò induce il rilascio dei batteri simbionti mediante defecazione o rigurgito. Dipoi si verifica la liberazione di una miscela di prodotti secretori/escretori, che includono piccole molecole, proteine e acidi nucleici. I componenti di questa mistura sono altamente tossici e si dislocano rapidamente, ragion per cui vengono indicati con il nome “venom proteins” (veleno di proteine).
In uno studio del 2017, pubblicato su Plos Pathongens, si è constatato che l’attivazione del nematode e la liberazione delle venom proteins non avvengono simultaneamente, bensì la quantità di molecole emesse dipende dalla tempistica e dal grado di attivazione. I ricercatori effettuarono l’attivazione di grandi gruppi di larve IJ e raccolsero i prodotti di secrezione/escrezione a diversi intervalli temporali (6 ore, 12 ore, 18 ore, 24 ore, 30 ore, 42 ore e 54 ore). Successivamente separarono le varie molecole mediante elettroforesi su gel e le visualizzarono tramite impregnazione cromoargentica (metodo di Golgi) (Fig. 4). Oltre a ciò, confrontarono la quantità di molecole prodotte dai nematodi associati con il batterio X. nematophila e i nematodi axenici (senza il batterio), scoprendo un profilo abbastanza simile nell’abbondanza delle proteine secrete (Fig. 4).

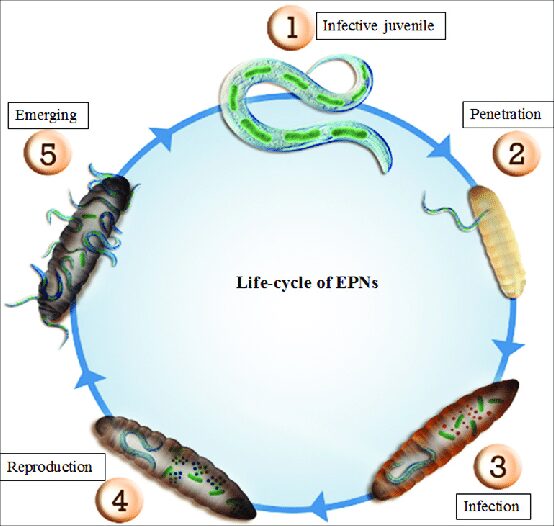
I batteri simbionti si espandono nell’emolinfa dell’insetto parassitato, il quale muore nel giro di due o tre giorni. I nematodi continuano a nutrirsi dei tessuti della vittima (che vengono liquefatti), si sviluppano in maschi e femmine, si accoppiano e procreano. A seconda della disponibilità delle risorse, nello stesso organismo ospite possono formarsi una o più generazioni e può avvenire la fuoriuscita di una grossa mole di larve immature infettive nell’ambiente, che vanno ad invadere altri ospiti e proseguono il loro ciclo biologico (Fig. 5).

S. carpocapsae, come anche gli altri nematodi entomopatogeni, può infettare diverse specie di insetti che vivono nel suolo, tra cui tarme, farfalle, coleotteri, scarabei, cavallette, grilli, piralidi, maggiolini e grillitalpa, sia allo stadio larvale che adulto.
Gli insetti uccisi dai nematodi della famiglia Steinermatidae sono caratterizzati da un colore marrone chiaro, mentre se vengono attaccati dai vermi della famiglia Heterorhabditidae diventano rossi e i tessuti assumono una consistenza gommosa.
La figura 6 riassume schematicamente il ciclo vitale dei vermi entomopatogeni.
Può capitare che il sistema immunitario della vittima riesca a difendersi dall’infezione, e su tale capacità giocano un ruolo importante le cellule chiamate emociti, che effettuano il processo dell’incapsulamento, ovvero, si legano alla superficie del nematode per formare una guaina di cellule sovrapposte. In aggiunta al sistema immunitario, gli insetti hanno sviluppato barriere comportamentali, morfologiche e fisiologiche verso i vermi entompatogeni. Tra quelle comportamentali abbiamo lo sfregamento della cuticola mediante le mandibole, allo scopo di rimuovere i nematodi attaccati alla superficie. Per quanto riguarda le strategie morfologiche, una di esse è il restringimento degli orifizi per limitare l’entrata dei vermi.
Identificazione
L’individuazione del nematode S. carpocapsae si fonda soprattutto su tecniche di biologia molecolare, e per affrontare tale argomento citiamo un lavoro pubblicato su Journal of Nematology nel 2020. In questo studio i ricercatori raccolsero diversi campioni di terreno nelle riserve di Toribìo e Tacueyò, nel Dipartimento di Cauca (Colombia). Le aree rurali scelte si trovavano tra 1757 e 2963 metri al di sopra del livello del mare, ed erano caratterizzate da temperature tra 13,9° C e 20° C. I campioni furono posti all’interno di scatole di plastica, e sulla superficie di ognuno furono aggiunte cinque larve di tarma maggiore della cera (Galleria mellonella) e cinque di tarma della farina (Tenebrio molitor). Le scatole vennero tenute al buio a 20° C per quattro giorni.
Le fasi successive furono la ricerca di larve morte uccise dai vermi entomopatogeni, l’isolamento delle larve immature infettanti e l’estrazione del DNA. In seguito gli scienziati effettuarono l’amplificazione per PCR (reazione a catena della DNA polimerasi) di un frammento della sequenza codificante per l’RNA ribosomiale 18S (18S rRNA), un frammento contenente le sequenze ITS1 (sequenza di DNA spaziatore interna di tipo 1), ITS2 (sequenza di DNA spaziatore interna di tipo 2) e 5.8S, e un frammento dell’rRNA 28S contenente le sequenze di espansione D2 e D3.
Le sei sequenze ottenute furono allineate e modificate mediante il software MEGA X, al fine di ottenere delle sequenze consenso (sequenze a cui si lega l’RNA polimerasi per iniziare la trascrizione); da queste ultime venne eseguita l’analisi filogenetica tramite il loro allineamento con sequenze di varie specie di nematodi entomopatogeni presenti nel database GenBank, usando il software MUSCLE (MUltiple Sequence Comparison by Log-Expectation).
Un articolo del 2006, pubblicato su Phytoparasitica, descrive un’ulteriore metodica, ovvero i polimorfismi di lunghezza dei frammenti di restrizione (RFLP), che si basa sull’utilizzo di particolari enzimi prodotti dai batteri, le endonucleasi di restrizione, che tagliano il DNA a livelli di zone caratteristiche, i siti di restrizione (Fig. 7). Questo metodo analitico venne realizzato sulla regione ITS del DNA ribosomiale di tre nematodi entomopatogeni (Steinernema feltiae, Steinernema carpocapsae e Heterorhabditis bacteriophora) isolati nella città di Ankara, in Turchia. Questa sequenza venne amplificata per PCR e digerita con nove enzimi di restrizione (Alu I, Dde I, Hae III, Hha I, Hind III, Hinf I, Hpa II, Rsa I e Sau 3AI). I frammenti risultanti furono poi separati mediante elettroforesi su gel d’agarosio.

Applicazioni in ambito agricolo
Come abbiamo accennato all’inizio, lo Steinernema carpocapsae si impiega come agente di controllo degli insetti che rovinano le coltivazioni, quindi come biopesticida.
È chiaro che i coltivatori non adotterebbero agenti biologici che non presentano un’efficacia paragonabile a quella dei tradizionali pesticidi chimici. I progressi tecnologici nella produzione, formulazione, controllo di qualità, tempi di applicazione dei nematodi entomopatogeni, e soprattutto la selezione ottimale degli habitat e degli insetti bersaglio, hanno ristretto il dislivello tra agenti chimici e nematodi. Di conseguenza, questi ultimi si sono dimostrati idonei nell’agricoltura e nell’orticoltura.
I vermi entomopatogeni sono notevolmente versatili contro gli insetti infestanti in diversi sistemi di coltivazione, anche se attualmente sono ancora poco adoperati. Come altri agenti biologici di controllo, i nematodi devono rimanere vivi e richiedono condizioni specifiche per essere validi. Infatti l’essiccamento o i raggi ultravioletti aboliscono la loro capacità insetticida; gli insetticidi chimici, al contrario, hanno meno limitazioni. Similarmente, i nematodi sono efficaci a un intervallo di temperatura più ristretto (20° C e 30° C) rispetto agli agenti chimici, e sono maggiormente influenzati dal tipo di suolo, dalla frequenza di irrigazione e dalla profondità del paglia, cioè lo strato di materia organica vegetale viva o morta che comprende radici, rizomi, steli e foglie (Fig. 8). Gli insetticidi basati sui nematodi possono essere inattivati se tenuti all’interno di veicoli caldi, non possono essere lasciati in taniche per lunghi periodi e sono incompatibili con molti prodotti chimici usati in agricoltura.
Anche i prodotti chimici hanno però diversi inconvenienti, tra cui tossicità, inquinamento e resistenza di alcuni insetti. L’implementazione accelerata dei nematodi all’interno dei sistemi di gestione integrata degli animali infestanti necessita di utenti più informati su come usarli in maniera idonea.

Considerando la biologia dei nematodi, le loro applicazioni dovrebbero prevedere degli accorgimenti per evitare la luce solare diretta, ad esempio utilizzandoli la mattina presto o la sera. Il suolo dell’area trattata dovrebbe essere mantenuto umido per almeno due settimane dopo aver collocato i nematodi. L’applicazione sopra il livello del suolo delle zone bersaglio è complicata a causa della sensibilità dei vermi alla disidratazione ai raggio ultravioletti. Tuttavia, sono stati raggiunti dei buoni risultati contro dei target che si trovano fuori terra, e di recente il miglioramento delle formulazioni ha incrementato gli approcci. In ogni caso, i nematodi devono essere applicati a una quantità sufficiente per uccidere gli insetti bersaglio; di solito occorrono 250000 larve immature infettanti per metro quadro della zona trattata. In aggiunta a ciò, è fondamentale scegliere le specie di nematodi appropriate a uno specifico insetto.
Grazie agli studi del dipartimento di Ricerca e Sviluppo di Koppert, si è giunti a una formulazione di Steinernema carpocapsae per il controllo biologico di una certa mole di insetti che abitano il suolo e che si nutrono di fogliame (Tabella 1). Questa è presente in commercio con il nome Capsanem (Fig. 9) in confezioni contenenti i nematodi in materiale di supporto inerte (86% S. carpocapsae e 14% materia inattiva).
| Lepidotteri | Coleotteri | Ditteri | Altri ordini |
| Larve di fillominatrice del pomodoro (Tuta absoluta) | Larve di dorifora della patata (Leptinotarsa decemlineata) | Larve di mosche scatelle (Scatella sp.) | Miridi (Nesidiocoris tenuis) |
| Larve di nottue (Spodoptera sp, Helicoverpa sp., Chrysodeixis chalcites, Agrotis sp., Autographa gamma, Duponchelia fovealis) | Larve di capnodio (Capnodis tenebrionis) | Larve di tipulidi/tipule dei prati (Tipula sp.) | Tingide del platano (Corythucha ciliata) |
| Larve di piralide del bosso (Cydalima perspectalis) | Grillitalpa (Gryllotalpa gryllotalpa, Grillitalpa americani Neoscapteriscus sp.) | ||
| Larve di Crambus hortuellus, Chrysoteuchia topiaria |

Fonti
- John Lucas, Mary Goetsch, Matt Fischer, Steven Forst. 2018. “Characterization of the pixB gene in Xenorhabdus nematophila and discovery of a new gene family”, Microbiology (Reading)
- Adler R. Dillman, John M. Chaston, Byron J. Adams, Todd A. Ciche, Heidi Goodrich-Blair, S. Patricia Stock and Paul W. Sternberg. 2012. “An Entomopathogenic Nematode by Any Other Name”, PLOS Pathogens
- Dihong Lu, Marissa Macchietto, Dennis Chang, Mirayana M. Barros, James Baldwin, Ali Mortazavi and Adler R. Dillman. 2017. “Activated entomopathogenic nematode infective juveniles release lethal venom proteins”, PLOS Pathogenes
- Laia Batalla-Carrera, Ana Morton, David Shapiro-Ilan, Michael R. Strand, and Fernando Garcìa-Del-Pino. 2014. “Infectivity of Steinernema carpocapsae and S. feltiae to larvae and adults of the hazelnut weevil, Curculio nucum: differential virulence and entry routes”, Journal of Nematology
- I. Alper Susurluk & Umut Toprak . 2006. “Molecular identification of three entomopathogenic nematodes from turkey by PCR-RFLP of the ITS regions”, Phytoparasitica
- Esteban Neira-Monsalve, Natalia Carolina Wilches-Ramírez, Wilson Terán, María del Pilar Márquez, Ana Teresa Mosquera-Espinosa and Adriana Sáenz-Aponte. 2020. “Isolation, identification, and pathogenicity of Steinernema carpocapsae and its bacterial symbiont in Cauca-Colombia”, Journal of Nematology