Abbiamo già visto la modalità con cui i batteri riescono a muoversi nel mezzo fluido grazie alla rotazione flagellare. In questo articolo tratteremo del fenomeno con cui i batteri decidono di dirigere i loro movimenti in dipendenza alla presenza di sostanze nutritive oppure di sostanze repellenti.
Il movimento flagellare
Mediante il movimento flagellare, un batterio può dirigersi verso ambenti ricchi di sostanze nutritive oppure allontanarsi da un ambiente in cui è contenuta una sostanza tossica.
Lo spostamento batterico da un ambiente povero di sostanze nutritive ad un ambiente che ne è ricco oppure l’allontanamento del batterio da un ambiente repellente è nominato chemiotassi (composta dalle parole greche χημεία, chemeia = chimica e τάξις, taxis = schieramento).
Il movimento di batteri dotati di flagello è un’alternanza tra gli spostamenti in linea retta (corse), che sono causati dalla rotazione dei flagelli in senso antiorario, e brevi soste durante le quali la cellula batterica, ruotando il flagello in senso orario, modifica l’orientamento (capriole). La descrizione appena fatta si riferisce al movimento della cellula batterica quando si trova in assenza di gradiente chimico. Cosa succede, invece, se il batterio deve fuggire da un ambiente in cui è presente una sostanza nociva oppure deve dirigersi da un ambiente povero di sostanze nutritive ad un ambiente più ricco? E, quindi, come riescono i batteri a “sentire” cosa è presente nell’ambiente in cui viaggiano? Scopriamolo insieme!
I sistemi chemosensoriali dei batteri
La superficie delle cellule batteriche è dotata di sistemi chemosensoriali che, durante il movimento cellulare, misurano in tempi diversi la concentrazione della specie chimica presente nel mezzo e, se necessario, ne valuta il cambiamento, riuscendo a trasdurre il segnale sentito in modo tale da modificare l’alternanza di corse e capriole. Dunque, se la cellula percepisce una diminuzione di una sostanza tossica oppure un aumento di una sostanza nutritiva, può inviare un segnale al motore del flagello che impedisce la rotazione del flagello in senso orario, diminuendo la probabilità di effettuare capriole e di cambiare orientamento. Per contro, se la cellula batterica percepisce una diminuzione delle sostanze nutritive oppure un aumento di sostanze tossiche, invia un segnale al motore flagellare che può ruotare in senso orario ed effettuare una capriola che permetta al batterio di modificare il proprio percorso ed allontanarsi dalla porzione del mezzo poco favorevole al suo nutrimento.

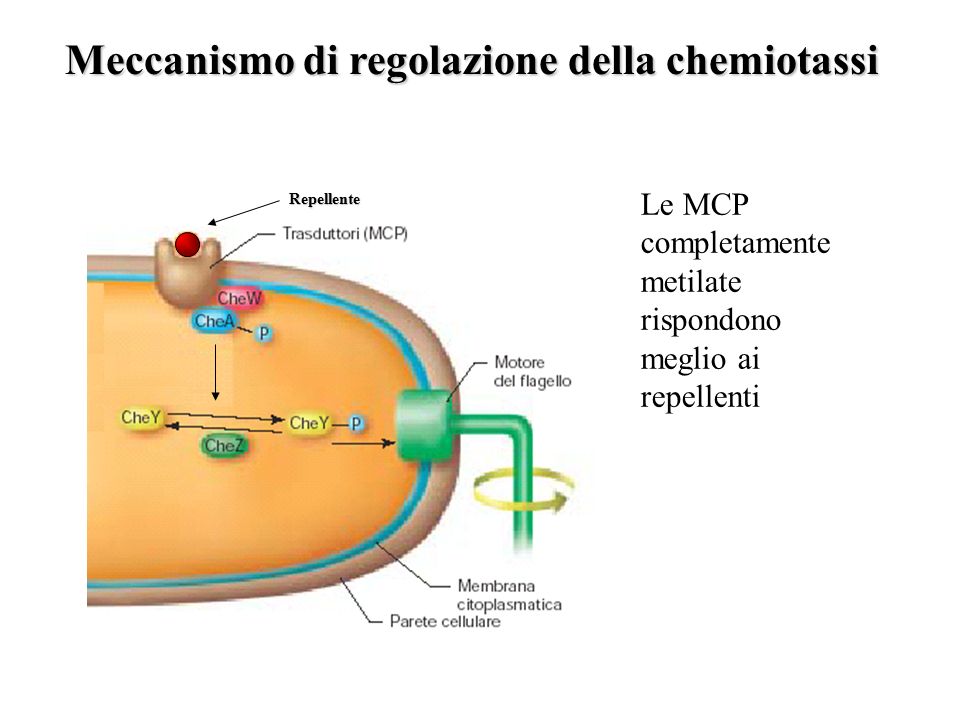
La via della segnalazione dei chemorecettori in E. coli
Sulla superficie del batterio Escherichia coli è stata rilevata la presenza di particolari chemorecettori chiamati proteine della chemiotassi metil-accettrici (MCP, methyl-accepting chemiotaxis protein). Tali proteine sono situate vicine ai poli sensoriali.
I chemorecettori MCP, quando metilati, si attivano e inducono i cambiamenti nella direzione dei movimenti attraverso il passaggio del segnale a recettori transmembranari e le proteine citoplasmatiche CheA e CheW.
Quando si verifica un aumento della sostanza nutritiva, la proteina CheA si attiva autofosforilandosi. La proteina CheA, attivata, fosforila, a sua volta, la proteina citoplasmatica CheY che interagisce con la proteina FliM del motore flagellare. L’interazione tra CheY e FliM promuove la rotazione del flagello in senso orario, causando cambiamenti nella direzione del percorso del flagello. La proteina CheB, invece, è una metile-esterasi che, quando fosforilata, è attivata. La sua attivazione determina la demetilazione dei residui di metil-glutamato presenti nelle proteine MCP e diminuendo l’attività chinasica di CheA.
Il processo di fosforilazione è altamente regolato. La proteina citoplasmatica CheZ è una fosfatasi che agisce su CheY fosforilata, assicurando una breve durata della rotazione in senso orario.
Infine, la proteina CheR compete con la proteina CheB nel controllo del grado di metilazione delle proteine MCP, metilando i residui di glutamato e aumentando l’attività chinasica di CheA.
Fonte
- Biologia dei microrganismi. Gianni Dehò, Enrica Galli. Casa editrice Ambrosiana. 2016, 72-73.
- Per un approfondimento sul movimento flagellare si rimanda a
http://www.microbiologiaitalia.it/2019/04/30/il-movimento-flagellare/
- Immagine in evidenza da https://www.igeasalute.com/escherichia-coli/
- Immagine 2 da https://slideplayer.it/slide/9176305/
Maria Chiara Langella