Pastorizzato, sterilizzato, microfiltrato, uperizzato. Il latte vaccino, quando crudo, è un alimento corposo e vivo, con tutti i suoi multiformi residenti batterici. Molti sono patogeni. Uno studio, oggi, valuta l’opportunità di regolarne il profilo microbiologico, mediante l’inaspettato supporto di altri batteri, nostri bio-alleati. Collaboratori di Purezza, si direbbe. Tutt’altro che pentiti.
I leggendari benefici nutrizionali di un alimento di crescita vero e proprio, come il latte crudo vaccino (figura 1), però, hanno un costo. Il rischio di incorrere in patogeni come lupi nascosti in un probiotico gregge, che proliferano e pullulano, anche in molti altri cibi.

Fonte: https://www.fisco7.it/2016/11/iva-al-4-sul-latte-fresco-solo-se-destinato-al-consumatore-finale/
C’è tutto un mondo, nel latte crudo
Da sempre, o quasi, pilastro dell’alimentazione umana, il latte, ed i suoi derivati, è un puntuale riferimento durante l’età evolutiva. Vitamina A e B12, macro- e micronutrienti, calcio a braccetto col sottostimato fosfato, fanno del latte, crudo in particolare, un ambìto concentrato biochimico. Ma non siamo i soli, Noi, genere umano, a riconoscerlo, e di certo non siamo i primi.
Il latte crudo, vaccino e non, è un habitat ideale per numerose forme microbiche. Oltre i batteri probiotici, infatti, dalla indubbia rilevanza, in termini di tecnologia alimentare, il microbiota lattico accoglie anche batteri deterioranti e patogeni, di ogni ordine e grado. Salmonella spp., Escherichia coli (STEC, ovvero Escherichia coli produttore della tossina Shiga), Listeria monocytogenes, Campylobacter spp. e Staphylococcus spp coagulasi-positivo. Per citarne alcuni.
Certo, la pastorizzazione ha risolto, da tempo, il nostro rapporto con questo materno alimento, eppure, oggi, cresce anche la curiosità, da moderni, per sapori antichi. Lo sapevano bene le aziende agrituristiche, di tutto il mondo. Almeno finché, l’evasione, naturalistica e non, è potuta essere non solo immaginata, come ormai da più di un anno. Dopotutto, il rischio di incorrere nella ingestione di patogeni non è perfettamente scongiurato, neppure con latte pastorizzato. Esistono ben le contaminazioni post-pastorizzazione, infatti, secondo Berge – Baars e colleghi.
I nemici del saveur crue
I temibili, per eccellenza, Listeria monocytogenes e Staphylococcus aureus, non indietreggiano neppure in refrigerazione. Come afferma lo studio recentissimo del gruppo guidato da Leclercq. Le comunità microbiche, residenti nella matrice lattea, e nei suoi prodotti di trasformazione, possono persino interferire le une sulle altre, interagendo, inibendosi, stimolandosi.
Lo scopo primo, di tutti i generi, è, chiaramente, l’accesso ai nutrienti, offerti dal mezzo di crescita, da cui dipende la loro esistenza e proliferazione. Per questo, essi mettono in campo composti antibatterici e prodotti metabolici; da condividere, se il caso apra a collaborazioni, invece che alla eliminazione del concorrente.
Ed è proprio in tali intellegibili manovre di supremazia esistenziale, che lo studio, freschissimo, di Julia Hahne ed André Lipski punta un fermo riflettore. Inutile dire, che si agogna una potenzialità antibatterica degli stessi batteri, da volgere a nostro vantaggio. Far di loro, stipiti pieni di risorse, dei reali collaboratori involontari o, addirittura, nolenti. Vorremmo tutto, come sempre. Il sapore di latte fresco crudo e la sicurezza microbiologica.
C’è da dire, che se ne studiano di modelli predittivi di rischio, su cibi e bevande animali, per veder di prevenire esiti patologici o addirittura mortali. Ma i modelli son fallaci, spesso, perchè poco tengono conto delle interazioni che orientano e dirigono fortune e sfortune dei ceppi residenti delle matrici alimentari.
Torneremo a bere latte crudo? Lo studio
Julia Hahne ed André Lipski sono partiti col selezionare isolati di specie, altamente abbondanti, nel latte crudo vaccino: 24, gli isolati, per la precisione. I generi sono: Acinetobacter, Pseudomonas, Streptococcus, Lactococcus, Escherichia, Chryseobacterium, Carnobacterium, Arthrobacter ed Hafnia. Tutti identificati grazie al profilo di acidi grassi ed alle sequenze geniche di rRNA 16S. Alla “carica dei 24”, poi, i ricercatori hanno affiancato i seguenti ceppi:
- L. monocytogenes DSM 20600T,
- S. aureus DSM 1104T,
- B. subtilis DSM 10T,
- M. luteus DSM 20030T.
Mentre i primi due sono facilmente associabili alle più perniciose infezioni alimentari, il Bacillus subtilis è prevalentemente responsabile del guasto, lipolitico e proteolitico, dei prodotti alimentari in cui si trovi ad alloggiare. Il ceppo Micrococcus, infine, farà, nelle fasi sperimentali, da sensore, sotto diverse sostanze antibatteriche, che compariranno nel medium di coltura.
Tutti i generi batterici, dei 24 isolati da latte crudo, sono cresciuti su TSA (Merck) a 30°C. Tutti, sì, tranne uno: Arthrobacter psychrolactophilus JZ RK-74, a 20°C, comprensibilmente psicrofilo. E, per spingere un po’ la crescita di L. monocytogenes DSM 20600T, aggiunta di estratto di lievito (6.0 g/L).
Cronache di laboratorio: il Drop Assay
I ricercatori hanno scandagliato subito le interazioni, intromissioni, ingerenze reciproche, tra ceppo e ceppo. Il Drop Assay ha, quindi, guidato le indagini analitiche: su TSA, supplementato con glucosio o lattosio, sempre a 0.25% v/w; distinto, per questo, in glc-TSA e lac-TSA.
Il saggio parte con un’incubazione overnight di ciascun ceppo, su TSB (Merck), ed aggiustamento della densità ottica a 625 nm, fino ad una concentrazione microbica di 0.1 CFU/mL. Diluizione 1:100 per i Gram-positivi e 1:1000 per i Gram-negativi.
Tale coltura cellulare viene poi seminata su piastra, mediante tampone sterile di cotone. Ed essa diviene, in realtà, il manto fondale che funge da indicatore, dei comportamenti batterici. Qui, i ricercatori hanno deposto spots, gocce di coltura da 10 uL, dei ceppi-test, ovvero dei 4 ceppi patogeni aggiuntivi. Quindi, segue incubazione delle piastre, a 30°C per 24 ore, ed a 10°C per 7 giorni. In modo, così, da ottenere crescite adeguate sia dei ceppi indicatori, sia dei ceppi-test. Il tutto, naturalmente, in duplicato.
Riferimenti visivi degli esiti si semplificano così: i ceppi-test sono inibitori di crescita, se, intorno agli spots, si creano zone d’inibizione (prive di cellule); i ceppi-test sono stimolatori di crescita, se, intorno agli spots, si evidenziano crescite cellulari, più che in altri punti della piastra (figura 2).
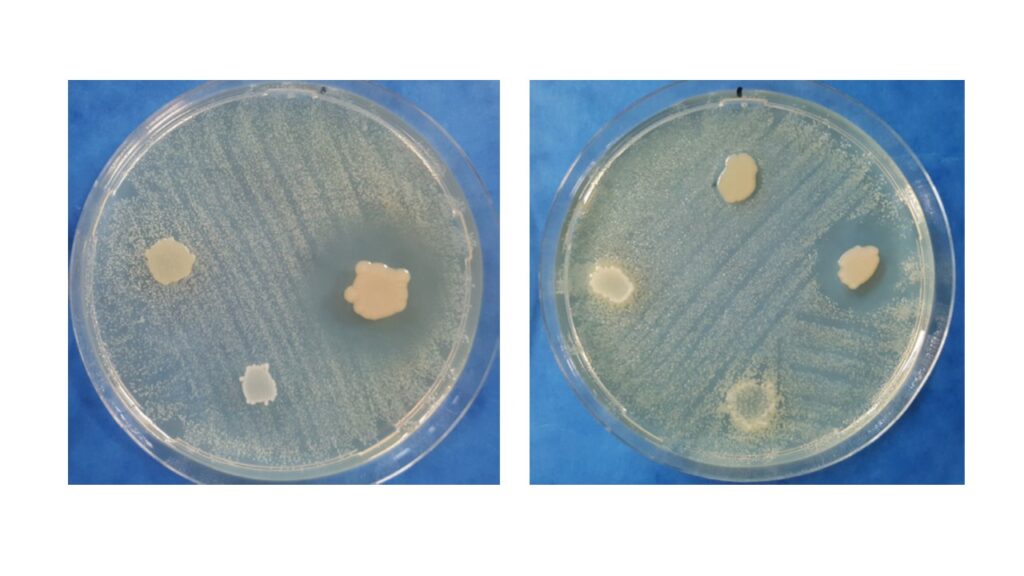
(a) Indicatore Listeriamonocytogenes DSM 20600T con gocce di ceppi-test Pseudomonas protegens JZ R-192 (1),
Carnobacterium maltaromaticum JZ RK-71 (2) e Acinetobacter johnsonii JZ RK-116 (3).
(b) Indicatore Ac. johnsonii JZ RK-116 con gicce di ceppi-test Pseudomonas veronii JZ RK-132 (1),
Ps. protegens JZ R-192 (2), Lact. lactis JZ R-198 (3) e Ca. maltaromaticum JZ RK-71 (4).
Fonte: https://sfamjournals.onlinelibrary.wiley.com/doi/abs/10.1111/jam.15056
Intanto, co-colture liquide
L’allestimento, contestuale, di co-colture liquide ha aperto nuovi scenari sui rapporti inter-microbici. I ricercatori hanno, infatti, valutato l’effetto del ceppo Pseudomonas protegens JZ R-192 su L. monocytogenes DSM 20600T e S. aureus DSM 1104T. E seguìto crescite combinate tra Carnobacterium maltaromaticum JZ RK-71 ed Acinetobacter johnsonii JZ RK-116.
Grosso modo, il 40% dei ceppi-test inibisce l’attività dell’indicatore piastrato, sia a 30°C che a 10°C. Inibitori di crescita, in particolare, sono: Lactococcus lactis JZ R-198, Ca. maltaromaticum JZ RK-71, Pseudomonas fragi JZ RK-32, Pseudomonas azotoformans JZ RK-64, Pseudomonas veronii JZ RK-132, Ps. veronii JZ R-189, Ps. protegens JZ R-192, Pseudomonas gessardii JZ R-150, Ps. gessardii JZ RK-44, Pseudomonas yamanorum JZ R-170, Pseudomonas helleri JZ RK-60, Pseudomonas synxantha JZ R-157, Pseudomonas fluorescens JZ R-133 e Pseudomonas arsenicoxydans JZ RK-27.
Individualità dirimenti, nel latte crudo
Nella singolarità ceppo-specifica, poi, 11 dei 14 stipiti di Pseudomonas valutati, mostrano capacità inibitoria delle crescite cellulari di patogeni, sia a 30°C che a 10°C. E colpiscono generi diversi dal proprio. Solo i ceppi Ps. protegens JZ R-192 e Ps. azotoformans JZ RK-64 mostrano attività inibitoria su altri Pseudomonas, prevalentemente a 10°C. A tutto vantaggio delle strategie potenziali di impiego, per sanificare cibi freschi, di là da venire.
Attitudini batteriche d’inibizione
Inoltre, le dimensioni delle zone d’inibizione, prodotte da Ps. protegens sui 4 ceppi-test, sono risultate doppie su lac-TSA, rispetto al glc-TSA. E quasi ogni ceppo differente dal genere Pseudomonas soffre l’interferenza, auspicabilissima, di Ps. protegens JZ R-192.
Stessi rassicuranti esiti sono, inoltre, confermati dalle curve di crescita delle co-colture liquide. Uno dei peggiori patogeni, L. monocytogenes DSM 20600T, per esempio, in co-coltura, decresce dopo 24 ore a 30°C, e dopo 48 ore a 10°C. Anche il numero massimo di cellule vitali risulta significativamente ridotto, in co-coltura, rispetto alla crescita in solitaria.
Identico decremento cellulare restituisce anche S. aureus DSM 1104T, in co-coltura con Ps. protegens JZ R-192, ma solo su TSB ed a 30°C. Esso decresce a 24 ore, ed a 114 ore non più del tutto rintracciabile. A 30°C, in prove su latte scremato, non è visibile, ed entrambi i ceppi, in realtà, decrescono dopo 24 ore. Qui, su latte scremato, ancora, nell’arco di 48 ore si registra un abbassamento del pH (da 7.1 a 5.1). In TSB normale, invece, le co-colture affrontano un innalzamento del pH (da 7.1 ad 8.1), nell’arco di 72 ore. Fino a pH 9.2, dopo 144 ore.
Attitudini batteriche di stimolazione
Il 30% dei ceppi-test hanno, invece, prodotto zone di stimolazione di crescita cellulare, intorno ai propri spots di deposizione, su piastra. Si tratta dei ceppi L. lactis JZ R-198, Ca. maltaromaticum JZ RK-71, H. paralvei JZ R-210 e S. parauberis JZ R-105 su Ac. johnsonii JZ RK-116, Ac. johnsonii JZ R-121 ed Ac. albensis JZ RK-19. Vale il viceversa.
In questo caso, a 30°C, le curve di crescita non descrivono un massimo della conta di cellule vitali. Anzi, entrambi i patogeni suddetti, su latte scremato, decrescono. Il pH scende da 7.1 a 4.9 nelle 48 ore. Tuttavia, in co-coltura, a 10°C, la conta di cellule vitali cresce sensibilmente, più che in solitaria.
Facoltà batteriche “extra”: i nostri bio-alleati contro i perfidi patogeni
Tutti i ceppi manifestano facoltà enzimatiche lipolitiche o proteolitiche a 30°C, come a 10°C. Ma l’83% di essi predilige i 30°C. Nella massa di popolazione, coinvolta nello studio, solo il genere Pseudomonas le possiede entrambe.
Ugualmente, per quanto concerne la duplice capacità, inibitoria/stimolante, del tessuto microbico della matrice “latte crudo”.
Pseudomonas, si conferma, dunque, un genere dalle avvincenti potenzialità antibatteriche, ad ampio spettro: mediante metaboliti secondari, tossine, siderofori. In particolare, Ps. protegens è un famoso batterio protettivo dei vegetali, che esprime prodotti atti al bio-controllo, come 2,4-diacetilfloroglucinolo e pioluteorina. Efficacissimi entrambi, contro ogni sorta di patogeno. Batteri, funghi, virus, nematodi.
I risultati dello studio, quindi, collimano con gli spunti rimasti in coda ai precedenti lavori sul tema. Che non sia solo la composizione qualitativa, della popolazione microbica, ma le trame che da essa si dipartono, a determinare la struttura di un alimento rotondo, come il latte crudo, non lo si può proprio più ignorare. Si riparta da qui.
Fonti
- https://sfamjournals.onlinelibrary.wiley.com/doi/epdf/10.1111/jam.15056
- Immagine d’anteprima fonte: https://pxhere.com/it/photo/627461
- Figura 1 fonte: https://www.fisco7.it/2016/11/iva-al-4-sul-latte-fresco-solo-se-destinato-al-consumatore-finale/
- Figura 2 fonte: https://sfamjournals.onlinelibrary.wiley.com/doi/epdf/10.1111/jam.15056
- https://www.microbiologiaitalia.it/batteriologia/patobiota-quando-la-simbiosi-si-fa-offensiva/
- https://www.sciencedirect.com/science/article/abs/pii/S074000202030071X
- https://www.frontiersin.org/articles/10.3389/fmicb.2020.01557/full?utm_source=F-AAE&utm_medium=EMLF&utm_campaign=MRK_1383094_51_Microb_20200721_arts_A
- https://www.frontiersin.org/articles/10.3389/fmicb.2020.01557/full?utm_source=F-AAE&utm_medium=EMLF&utm_campaign=MRK_1383094_51_Microb_20200721_arts_A
Il lavoro che la D.ssa Giuliani ha cortesemente riproposto pone l’accento su come, alle volte, possa essere incompleta la visione del mondo microbiologico che ci circonda.
Spesso infatti, l’approfondita conoscenza di un determinato gruppo microbico, ad esempio i batteri lattici o le pseudomonadi, tanto per rimanere nell’ambiente latte, possa essere d’ostacolo a vedere oltre.
Nel caso degli Pseudomonas, essi sono ben conosciuti, se non temuti, nel mondo dei prodotti lattiero-caseari proprio in ragione del loro psicrofilia che, combinata con un arsenale enzimatico che interessa centinaia di molecole organiche, è causa delle alterazioni nutrizionali ed organolettiche dei prodotti freschi.
Tuttavia le relazioni tra le varie popolazioni del microbioma del latte sono molto più estese e sconosciute di quanto oggi è dato di sapere. In tale direzione si muove l’articolo di Hahne e Lipski che dimostra un nuovo, e potenziale, ruolo di ceppi di Pseudomonas nell’equilibrio tra le varie popolazioni microbiche naturalmente presenti nel latte crudo.
Il proseguire degli studi verso strade differenti da quelle normalmente battute, ovvero vedere Pseudomonas come principale genere alterativo di latte e formaggi freschi, potrebbe portare ad rivalutare, o quantomeno ad ampliare, l’importanza di tale genere nella contaminazione del latte e prodotti derivati. E forse anche a “sfruttare” alcune loro capacità per il miglioramento della qualità organolettica, nutrizionale e salutistica degli alimenti.