Il quorum sensing, letteralmente “misurazione della quantità di individui che compone una società di batteri”, è un meccanismo di comunicazione cellula-cellula utilizzato dalla maggior parte dei batteri Gram-positivi e Gram-negativi e basato sul controllo della densità di popolazione. Ma cosa ha a che fare con la sincronizzazione delle onde cerebrali?
Le cellule batteriche producono in modo costitutivo specifiche molecole segnale con la funzione di trasmettere il messaggio ad altri batteri in grado di recepirlo. Queste molecole prendono il nome di autoinduttori e contengono informazioni utili per la sopravvivenza, come quelle riguardanti, ad esempio, le condizioni dell’ambiente circostante. L’autoinduttore diffonde liberamente attraverso la membrana cellulare ma solo ad elevate concentrazioni s’innesca il meccanismo che consente ai batteri di comunicare.
Scambio di informazioni
Che i batteri fossero in grado di comunicare tra loro non è certo la notizia dell’anno. Difatti già a partire dal 1960 circa, la comunità scientifica fu in grado di descrivere il sistema del quorum sensing, utilizzato dalle cellule batteriche di Vibrio fischeri per scambiarsi informazioni. Il meccanismo è stato scoperto casualmente, osservando il fenomeno di bioluminescenza nel batterio Vibrio fischeri (Figura 1), in grado di emettere luce solo quando il numero di cellule batteriche che compone la popolazione è elevato.
Perchè, per la precisione, trattasi di un sistema di regolazione trascrizionale, dipendente dalla densità cellulare, che consta di due elementi: la molecola segnale contenente l’informazione e definita “autoinduttore” ed un regolatore trascrizionale. I batteri preferiscono vivere in aggregati cellulari e periodicamente “tastano il terreno” circostante valutandone le condizioni ambientali; per questo producono in modo costitutivo specifici segnali chimici che possono essere captati da altri batteri (solitamente appartenenti alla stessa specie e facenti parte della stessa popolazione) tramite recettori extracellulari, permettendo così una comunicazione cellula-cellula.
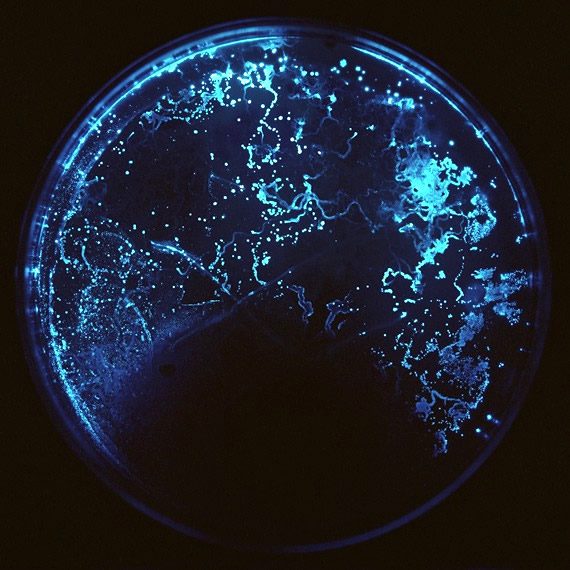
Figura 1. Fenomeno della bioluminescenza in piastra Petri. Alcuni organismi viventi sono in grado di convertire l’energia chimica in energia luminosa; il fenomeno è conosciuto col nome di bioluminescenza ed è diffuso soprattutto tra gli organismi marini dotati di appositi organi in grado di produrre luce: i fotofori. Anche alcuni insetti, e l’esempio più lampante (permettetemi il gioco di parole) è quello delle lucciole, sono in grado di produrre energia luminosa ma sembra che i meccanismi alla base dell’emissione di luce da parte di differenti organismi viventi siano vari, così come le ragioni evolutive che hanno portato allo sviluppo di questa capacità in maniera indipendente nei diversi gruppi tassonomici. Nell’immagine è rappresentata la bioluminescenza, osservata in vitro, emessa dal batterio Vibrio fischeri. Il fenomeno non potrebbe essere possibile in assenza di appositi geni, un sistema enzimatico adeguato e la capacità delle cellule batteriche di comunicare tra loro. Vibro fischeri è stato per molto tempo oggetto di studio, al fine di comprendere meglio la bioluminescenza microbica ed il quorum sensing. La ricerca su questo batterio Gram-negativo ha permesso di individuare il significato funzionale di entrambi gli eventi: Vibro fisheri è presente prevalentemente in habitat marini dove è in grado di instaurare rapporti simbiontici batterico-animale favorevoli alla propria sopravvivenza. La simbiosi con il cefalopode Euprymna scopoles è una delle associazioni più forti: il calamaro fornisce sostanze nutritive al batterio che, in cambio, lo protegge dai predatori permettendone il camuffamento tramite il fenomeno della controilluminazione. L’intera comunità di Vibro fisheri, nascosta nel fotoforo dell’animale, in seguito alla comunicazione intercellulare, emette energia luminosa in grado di nascondere la sagoma del cefalopode.
Geneticamente parlando
Quando la densità di popolazione è bassa l’autoinduttore diffonde semplicemente all’esterno della cellula; nello spazio extracellulare si lega al recettore specifico presente sulla membrana cellulare per promuovere lo scambio di informazioni. A sua volta la cellula batterica interessata sintetizzerà una nuova molecola “autoinduttore” che verrà rilasciata nello spazio extracellulare, pronta a legarsi al recettore di un’altra cellula batterica.
Ma cosa succede quando la densità di popolazione aumenta? Ebbene, quando accresce il numero di cellule che compongono l’aggregato batterico, di conseguenza aumenta anche la concentrazione di molecole segnale che si accumula all’esterno della cellula. Naturalmente la molecola si muoverà secondo gradiente di concentrazione verso l’interno della cellula e qui l’autoinduttore si legherà al regolatore trascrizionale promuovendo o reprimendo l’espressione di geni bersaglio: a seconda che si tratti di attivatore o repressore trascrizionale si potrà determinare l’attivazione o lo spegnimento di specifici processi metabolici e/o cellulari.
L’attivazione del quorum sensing presuppone il raggiungimento di un livello soglia da parte della popolazione batterica, inteso come numero minimo di cellule, per poter gestire una certa quantità di autoinduttori in grado di determinare poi un effetto “a cascata”. La sintesi di queste molecole è dunque associato al biofilm.
Le nuove frontiere della scienza
L’emissione di luce, per il batterio in questione, non rappresenta altro che un meccanismo per mettersi in evidenza, per rendere visibile l’intera popolazione. Si potrebbe perciò pensare che l’attività di migliaia di batteri sia coordinata ma, in virtù di quanto già detto in precedenza riguardo il meccanismo di funzionamento delle molecole segnale, sappiamo che questo non è vero.
Tuttavia l’attività delle cellule può essere coordinata artificialmente tramite l’ingegneria genetica: in un esperimento condotto all’Università della California un team di ricercatori ha sviluppato una simile capacità in un ceppo di Escherichia coli ingegnerizzato. Naturalmente, a differenza di Vibrio fischeri, Escherichia coli non produce alcuna proteina fluorescente ma, per esigenze di laboratorio, il batterio è stato modificato per esprimere tre proteine in risposta ad un autoinduttore chiamato AHL: un enzima che consente un aumento della sintesi di AHL, una proteina fluorescente ed un inibitore trascrizionale in grado di arrestare l’espressione genica di tutte e tre le proteine al raggiungimento di un valore soglia della concentrazione di AHL.
Venne notato che a basse densità di popolazione i batteri producevano AHL, ma nessuna delle altre tre proteine. A densità più elevate invece, AHL iniziava ad accumularsi all’interno delle cellule innescando un meccanismo di feedback positivo: i batteri sintetizzavano le tre proteine per cui erano stati progettati e dunque anche l’enzima che consentiva una più rapida ed efficace sintesi di AHL. Ciò ha permesso di osservare il fenomeno della bioluminescenza nella colonia di batteri fino a quando non si è accumulata una concentrazione sufficiente di AHL nello spazio extracellulare per formare una riserva di molecole segnale in grado di legarsi contemporaneamente a più recettori presenti sulla membrana di diverse cellule batteriche: l’emissione di luce si era sincronizzata e poteva essere interrotta solo al raggiungimento di un nuovo valore soglia della concentrazione di AHL grazie alla presenza dell’inibitore trascrizionale.
E se invece di emettere luce somministrassero farmaci?
La sincronizzazione dell’attività di migliaia di batteri potrebbe rivelarsi molto utile a scopi medici: pensiamo alla possibilità di utilizzare, all’interno dell’organismo umano, una colonia di cellule batteriche ingegnerizzate allo scopo di erogare dosi precise di farmaci o ormoni in momenti specifici della giornata, ad esempio cellule batteriche che rilasciano contemporaneamente impulsi di insulina in risposta ai cambiamenti della concentrazione di zucchero nel sangue ed in grado di arrestarne poi la sintesi al raggiungimento di un valore soglia, grazie all’inibitore trascrizionale.
Oltre ad un nuovo modo di somministrare farmaci, il controllo del quorum sensing potrebbe aprire nuove strade e portare ad una conoscenza più approfondita sulle capacità di apprendimento e di memorizzazione e sul sonno, specialmente sui disturbi correlati a questi eventi, che si pensa si verifichino quando la sincronizzazione tra i neuroni è anormale.
Un esempio pratico: neurofisiologia del sonno
Il sonno è una condizione fisiologica necessaria che si presenta con aspetti diversi in tutti gli animali. Si tratta di un processo attivo che per convenzione viene suddiviso in 2 fasi e 5 stadi consecutivi ciclici e che coinvolge diverse componenti anatomiche cerebrali; tra queste ricordiamo l’ippocampo, le aree corticali temporo-occipitali, il nucleo soprachiasmatico, il circuito limbico ed alcuni nuclei talamici. Essendo il tessuto nervoso caratterizzato costantemente da un’attività elettrica, è possibile andare a rilevare, tramite l’elettroencefalogramma (EEG), onde elettromagnetiche peculiari, sia nella fase di sonno che in quella di veglia.
Queste onde ritmiche e ripetitive hanno diverse caratteristiche e vanno dall’avere bassa ampiezza (μV) ed alta frequenza (Hz), all’avere bassa frequenza e grande ampiezza (Figura 2).

Figura 2. Frequenza ed ampiezza delle diverse onde cerebrali.
Ritmi cerebrali
Le onde α e le onde β sono caratteristiche dello stato di veglia, in cui si registra un’attività EEGrafica corticale desincronizzata: in particolar modo le onde β sono tipiche di un’intensa attività mentale mentre le onde α si possono riscontrare in condizioni di riposo ad occhi chiusi, in uno stato meditativo e durante la fase dell’addormentamento.
Quando dallo stato di veglia si passa ad uno stato di sonnolenza, ovvero lo stadio 1 del sonno, la frequenza dell’attività cerebrale varia, le onde α si sostituiscono gradualmente alle onde onde θ e l’attività EEGrafica diventa sincrona. La sincronizzazione determina la presenza esclusiva di onde nella banda θ/δ con conseguente alterazione dello stato di coscienza e ridotta risposta agli stimoli sensoriali.
Dopo essersi addormentato, il soggetto passa progressivamente dallo stadio 1 del sonno allo stadio 2, caratterizzato sempre dalle onde θ e dalla presenza di onde elettroencefalografiche ad alto voltaggio: le sequenze SPINDLE ed i complessi κ.
Si passa poi allo stadio 3 e 4 del sonno, dove invece sono presenti solo le onde δ. In poche parole, nel procedere attraverso le fasi 2, 3 e 4, il sonno si fa più profondo e la frequenza dell’attività encefalografica diminuisce sempre di più (Figura 3).
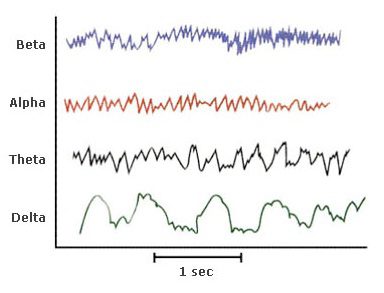
Figura 3. Rappresentazione grafica delle diverse onde cerebrali. Tracciati dell’attività elettrica ritmica del cervello ottenuta tramite registrazione poligrafica dell’elettroencefalogramma.
Le fasi del sonno
Questi primi 4 stadi definiscono la fase NON-REM dello stato di sonno, caratterizzata da un’architettura completamente differente rispetto alla fase REM, stadio 5 del sonno, che segue subito dopo. Il sonno N-REM è definito anche sonno ortodosso ed è caratterizzato da un rallentamento metabolico che non si registra invece nella fase REM del sonno.
Un’ulteriore differenza molto marcata tra le due fasi del sonno è la capacità di sognare, che si registra prevalentemente nella fase REM e non nella fase N-REM. La durata di ciascuno stadio è variabile ma in ogni individuo adulto l’intero ciclo (alternanza fase NREM-fase REM) si ripete circa ogni 90 minuti (Figura 4). Il sonno REM è conosciuto anche come sonno paradosso e rappresenta una fase del ciclo del sonno molto particolare poiché ricorda lo stato di veglia o lo stadio 1 del sonno NON-REM: si registra difatti un EEG con frequenze miste per cui è facile intuire che l’attività EEGrafica a livello corticale è desincronizzata, sono presenti movimenti oculari rapidi (REM dall’inglese, acronimo che sta per rapid eye movements), un basso tono muscolare per inibizione degli α e γ motoneuroni onde evitare di mimare nella realtà ciò che accade nel sogno e vi è alto consumo di ossigeno da parte delle cellule.
L’oscillatore talamo-corticale
A livello neurofisiologico le transizioni veglia-sonno e sonno-veglia sono controllate dall’adeguato funzionamento di un circuito cerebrale ben definito: il circuito talamo-corticale che genera un’attività autoritmica data da oscillazioni di onde lente e sincrone nella banda θ/δ in alcune condizioni funzionali, la più importante delle quali è sicuramente il sonno profondo.
L’attività è dunque periodica ma coincidente, cosa che suggerisce l’esistenza di neuroni che tendono ad assumere un comportamento collettivo scaricando potenziali d’azione in maniera simultanea. Durante il sonno profondo dunque il cervello entra in una fase di ipersincronismo che interessa l’intera corteccia cerebrale ed il lavoro dei neuroni può aumentare di circa il 10% (in poche parole l’organismo riposa e recupera energia ed il cervello è sempre vigile ed operoso).
In alcuni soggetti (umani ed animali) è stata osservata una mancata sincronizzazione dell’attività cerebrale durante e dopo la fase dell’addormentamento, con conseguente immediata insorgenza dei più comuni disordini del sonno: dissonnie (insonnia, ipersonnia, narcolessia, cataplessia) e parasonnie (sonnambulismo, bruxismo).
Grazie alla polisonnografia, esame che registra l’attività elettroencefalografica mentre il soggetto dorme, è possibile osservare come ed in quale fase, l’architettura dell’intero ciclo del sonno viene modificata. Vengono poi valutati altri parametri come i movimenti oculari e muscolari, la frequenza respiratoria e quella cardiaca, il flusso ematico cerebrale ed il tono muscolare antigravitario, al fine di individuare la causa e la corretta patologia del sonno.
A seconda della categoria di appartenenza difatti (dissonnie o parasonnie), esistono terapie farmacologiche appropriate che non sempre sono sufficienti per debellare il problema.

Figura 4. Ciclo del sonno della durata fisiologica di circa 90-120 minuti in un adulto sano.
I disordini del sonno
La cataplessia è un grave disturbo caratterizzato da fulminee incursioni di sonno REM durante la veglia. Il paziente cataplettico che prova o ricorda forti emozioni (felicità, tristezza, rabbia) è soggetto ad un’improvvisa atonia posturale, per cui si riversa a terra, ed è colpito da paralisi a causa della perdita del tono muscolare. Questi attacchi cataplettici possono essere più o meno gravi ed hanno una durata variabile (da pochi minuti a qualche ora). Nei casi documentati più seri il soggetto è stato dichiarato morto per poi rialzarsi in obitorio: la pressione arteriosa si è abbassata talmente tanto che, assieme ad un rallentamento nella frequenza cardiaca, il paziente sembrava clinicamente morto. Durante tutto la durata dell’attacco cataplettico il soggetto rimane cosciente e ad occhi aperti è in grado di vedere ciò che succede attorno a sé ma non è in grado di interagire con l’ambiente circostante.
Tra i disordini del sonno più diffusi vi è l’insonnia, cui, erroneamente, non vengono associate spiacevoli conseguenze, almeno non severe quanto nel caso della cataplessia. Esperimenti condotti con animali da laboratorio, hanno però dimostrato che la privazione di sonno non è compatibile con la vita: tra le prime conseguenze dell’assenza di riposo vi è l’alterazione delle funzioni fisiologiche gestite dal Sistema Nervoso Autonomo come conseguenza del danno alle strutture cerebrali sottocorticali.
Il sistema ipotalamico è il primo ad essere colpito: viene meno la termoregolazione del soggetto, cui seguono reazioni più istintive come l’autolesionismo e la mancata cura dell’igiene personale, l’aumento incontrollato dell’assunzione di cibo ma una smisurata perdita di peso ed infine la morte entro due settimane.
Un processo attivo
Il sonno dunque non è affatto un fenomeno accessorio, anzi, la funzione ristorativa ad esso associato è di vitale importanza per la sopravvivenza di tutte le specie viventi. Poter risolvere, o quantomeno limitare i danni associati alle patologie del sonno è possibile grazie alla sincronizzazione dell’attività dei neuroni talamici e corticali, ed in questo ci sono di grande aiuto i microrganismi.
Carla Caianiello
Fonti:
- Danino T., Mondragòn-Palomino O., Tsimring L., Hasty J. (2010) A synchronized quorum of genetic clocks. Nature; 463(7279), 326–330. doi: 10.1038/nature08753.
- https://it.wikipedia.org/wiki/Bioluminescenza
- https://www.microbiologiaitalia.it/2018/05/08/il-genoma-di-vibrio-fischeri-bioluminescenza-quorum-sensing-e-spunto-per-nuove-ricerche/
- https://www.newscientist.com/article/dn18418-bacteria-rewired-to-flash-in-sync
- https://static.medicitalia.it/public/uploadedfiles/minforma/maurocolangelo_fasi-del-sonno.jpg
- http://www.thecymartist.com/battiti-binaurali
- Kandel E.R., Schwarts J.H., Jessel T.M. Principi di neuroscienze. Casa Editrice Ambrosiana (2003). Terza Edizione
- Reading N.C., Sperandio V. (2006) Quorum sensing: the many languages of bacteria. FEMS Microbiology Letters; 254(1):1-11. doi: 10.1111/j.1574-6968.2005.00001.x
{kind=link}