Soltanto i neuroni e le cellule del muscolo scheletrico e cardiaco sono capaci di generare un segnale elettrico che si propaga sulle lunghe distanze. Per questo motivo sono definite cellule eccitabili, mentre il segnale è detto potenziale d’azione, con significato di trasmissione di informazioni.
Basi del potenziale d’azione (PdA): potenziale di membrana di riposo
Circuiti RC e legge di Ohm
Per introdurre il concetto di Potenziale d’azione (PdA), introduciamo prima quello del potenziale di riposo di membrana. Torniamo sui banchi di scuola e rispolveriamo un concetto che qualcuno potrebbe già aver sentito: i circuiti RC, ma cosa c’entrano? Immaginiamo una membrana cellulare fatta dal doppio strato fosfolipidico idrofobico (dunque un isolante) che separa un ambiente intracellulare e uno extracellulare, dove da una parte e dall’altra ci sono ioni che vogliono attraversarla. A cavallo della membrana ci sono due motori (detti driving force) che guidano la direzione degli ioni:
- Gradiente chimico (E): dato dalle concentrazioni degli ioni tra i due ambienti transmembranari, rivolto dall’ambiente più concentrato a quello meno concentrato;
- Gradiente elettrico (o potenziale di membrana) (V): le cellule eccitabili a riposo hanno un potenziale negativo all’interno con valore di -70mV.
Dalla combinazione dei due deriva il gradiente elettrochimico che guida il flusso ionico attraverso la membrana. Tuttavia, poiché gli ioni hanno una carica (+ o -), non possono liberamente attraversare la membrana, lo fanno solo in corrispondenza dei rispettivi canali ionici, dispersi qua e là nella membrana, ma che possono essere aperti o chiusi.
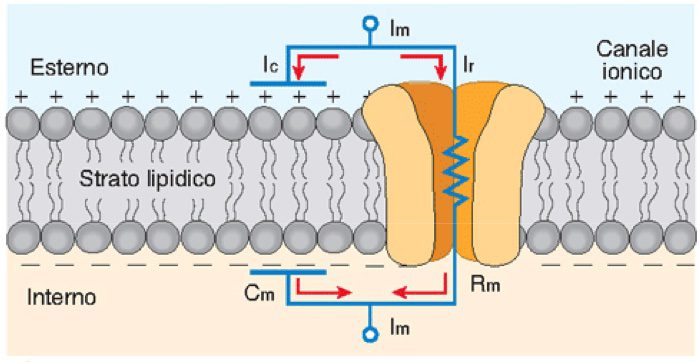
Abbiamo allora tutti gli elementi di un circuito elettrico RC (figura 1):
- i = corrente ionica;
- ∆V, o meglio E-V = gradiente elettrochimico che muove il flusso ionico;
- g = conduttanza ionica, calcolata come il reciproco della resistenza del circuito (1/R), che è un indice di quanto i canali ionici sono aperti o chiusi.
Secondo la legge di Ohm i = g(E-V), tanto più g è alto (ovvero i canali sono aperti) o tanto più il gradiente elettrochimico è alto, e tanto maggiore è il flusso ionico che genera una corrente transmembranaria.
Quali ioni sono coinvolti?
Per semplicità consideriamo che gli attori principali del PdA sono due: sodio (Na) e potassio (K). Il Na è molto più concentrato in ambiente extracellulare, mentre il K in quello intracellulare. Per comprendere come ciascuno ione si muove attraverso la membrana è stata messa a punto un’equazione, detta equazione di Nerst. Questa permette di calcolare che quella differenza di potenziale è capace di contrastare il loro naturale gradiente chimico, e arrivare all’equilibrio elettrochimico. Da questa equazione si ottiene che il potenziale di equilibrio di K è circa -100mV. Quello del Na è positivo, circa +40mV (da testo a testo i valori possono cambiare, è importante osservare le grandezze).
Poiché il potenziale di membrana a riposo è negativo (-70mV) la situazione di membrana a riposo è molto più vicina all’equilibrio del K anziché del Na. Dunque, i canali al K sono praticamente tutti aperti, e la membrana è virtualmente permeabile a questo ione instaurando correnti ioniche passive. Dunque, i canali al K sono detti canali di perdita (o di leak o background) e sono del tutto prevedibili con la legge di Ohm.
Da notare che il potenziale di riposo della membrana è leggermente meno negativo di quello di equilibrio del K (-70mv vs -100mV circa). Ciò indica che non c’è solo il K, bensì esiste un equilibrio multi-ionico (con anche calcio, cloro e altri ioni) che genera il potenziale di riposo di -70mV. Questo è calcolabile con l’equazione di Goldmann che tiene conto di tutti gli ioni presenti (l’equazione di Nerst considera una specie ionica e basta). Tale equazione dipende da 3 fattori: polarità della carica di ciascuno ione, permeabilità della membrana a ciascuno di essi, concentrazioni intra ed extracellulari.
Da correnti passive a correnti attive: potenziale d’azione
Abbiamo visto come a riposo la membrana ha una permeabilità del K molto maggiore rispetto a quella del Na (di quasi 100 volte). Gran parte dei canali al K sono aperti, mentre quelli del Na sono chiusi. Tuttavia, se ci si discosta dal potenziale di membrana di riposo di -70mV la situazione cambia:
- iperpolarizzazione: il potenziale di membrana diventa ancora più negativo (< -70mV) e i canali al Na si chiudono ancora di più, mentre quelli del K si aprono maggiormente. La corrente risulta essere uscente e fatta da K.
- depolarizzazione: il potenziale di membrana diventa meno negativo fino a diventare positivo (> 0mV). Avvicinandosi al potenziale di equilibrio del Na (di circa +40mV), i canali del K si chiudono e si aprono quelli del Na. Nel momento in cui il potenziale di membrana diventa sempre meno negativo si arriva a un valore, detto valore soglia, in cui la corrente che si genera non è più prevedibile dalla legge di Ohm. Da fenomeno passivo diventa un fenomeno attivo. La corrente risulta essere entrante e fatta da Na.
Dunque, il PdA si genera sulla base di modificazioni di rapporti di conduttanza di membrana per lo più per gli ioni K e Na. A riposo la permeabilità dello ione K è quasi 100 volte maggiore di quella del Na. La situazione si ribalta al raggiungimento del valore soglia e con il conseguente innesco del PdA.
Il PdA risulta quindi essere una rapida e transitoria depolarizzazione di membrana. E’ definito come un fenomeno del tipo tutto o nulla ed è detto stereotipo. Ciò significa che nel momento in cui si innesca, ovvero se si supera il valore soglia, non è modulabile in intensità, bensì raggiungerà sempre lo stesso valore di depolarizzazione, molto simile al potenziale di equilibrio dello ione Na, ovvero circa +45mV. Inoltre, il PdA è autorigenerativo propagandosi da una cellula all’altra senza attenuarsi.
PdA nelle fibre nervose e muscolari scheletriche
Il PdA dura dai 2 ms (millisecondi) nelle fibre nervose a 5-6 ms nel muscolo striato.

- Membrana a riposo: il potenziale di membrana è -70mV, assimilabile a una membrana permeabile allo ione potassio.
- Depolarizzazione lenta: a seguito di uno specifico stimolo, la membrana diventa permeabile allo ione sodio a causa dell’apertura dei canali al sodio voltaggio-dipendenti. Si innescano correnti entranti che depolarizzano la membrana.
- Depolarizzazione rapida: Raggiunto il valore soglia, si innesca un fenomeno “esplosivo” che fa aprire tanti altri canali al sodio. Il potenziale di membrana arriva a valori vicini al potenziale di equilibrio del Na (+45mV), e viene detta fase di eccedenza (o overshot).
- Ripolarizzazione: raggiunto l’apice di depolarizzazione, i canali al sodio tornano a chiudersi, mentre si aprono quelli al potassio. Correnti uscenti di potassio riportano la membrana a valori di potenziale negativi.
- iperpolarizzazione postuma (o after-potential): i canali al potassio sono aperti, quelli al sodio chiusi, ciò comporta un abbassamento del potenziale di membrana al di sotto del valore di -70mV, vicino al potenziale di equilibrio dello ione K (circa -100mV).
Da notare, inoltre, che i canali al sodio sono voltaggio dipendenti, ovvero si aprono man mano che il potenziale di membrana si sposta verso valori positivi (o meno negativi di -70mV). Ciò significa che durante la fase di depolarizzazione lenta l’apertura dei canali al Na comporta un ingresso nella cellula di questo ione, comportando depolarizzazione di membrana che fa aprire a sua volta altri canali al Na voltaggio-dipendenti. Si dice che l’effetto è rigenerativo, ovvero si autoalimenta. Questo fenomeno si chiama ciclo di Hodgkin (sistema a feedback positivo). Altra proprietà dei canali al sodio è che possono essere inibiti con la tetradotossina (TTX), un veleno estratto dal pesce palla.

Canali al sodio voltaggio-dipendenti (ingresso di sodio)
Questo canale è fatto da una proteina divisa in 4 domini, ciascuno composto da 6 alfa-eliche trans-membrana con estremità amino- (NH2) e carbossi- (COOH) terminali intracellulari. Questa è la struttura detta pore forming determinando la formazione del canale che permette il passaggio di ioni Na+ attraverso la membrana idrofobica. Ad oggi sono note 9 isoforme che variano per le strutture accessorie oltre al poro centrale, e prendono in nome da 1.1 a 1.9.
Come si osserva in figura 4, l’ansa S4 di ciascun dominio corrisponde al sensore di voltaggio. Sempre riferendoci alla figura, in azzurro e in giallo sono indicati rispettivamente il sito di inattivazione e il cancello di inattivazione. La selettività del canale al sodio è data da una porzione di ogni dominio detta ansa P che costituisce un filtro ionico lasciando passare solo lo ione di interesse.

I meccanismi che mediano l’attivazione e l’inattivazione del canale al sodio sono la porta di attivazione e il cancello di inattivazione. La prima a è una porta voltaggio dipendente, la seconda è una porta “temporizzata”. Mentre il potenziale di membrana è a riposo il cancello di inattivazione è aperto, mentre la porta di attivazione è chiusa. Tuttavia è capace di aprirsi se il potenziale si depolarizza, segnale che lo fa aprire rendendo il canale pervio per gli ioni Na.
Con il permanere della depolarizzazione il cancello di inattivazione si chiude in modo tempo dipendente. Tale chiusura avviene nella fase finale di depolarizzazione innescando il periodo di refrattarietà. Esso di suddivide prima in refrattarietà assoluta (nessun’ulteriore depolarizzazione ne consentirebbe l’apertura) e poi relativa (solo una depolarizzazione molto sopra soglia lo attiverebbe). Una conseguenza della refrattarietà è che la propagazione del potenziale d’azione in un tessuto eccitabile è unidirezionale.
Canali al potassio di leakage (uscita di potassio)
I canali al K sono anche detti di perdita, o di background o di leakage perché costitutivamente attivi contribuendo a stabilizzare il potenziale di membrana di riposo ai valori di -70mV (vicini al potenziale di equilibrio del potassio). Sono formati da dimeri di subunità alfa, ciascuna fatta da 4 segmenti transmembrana (S1-S4). Le regioni extracellulari tra i segmenti S1-S2 e S3-S4 costituiscono le anse P, che come visto per i canali al sodio, sono importanti per la selettività ionica. Esistono 15 membri della famiglia di questi canali, che mediano correnti sempre al potassio, ma con caratteristiche diverse come vedremo poco più avanti nel descrivere il potenziale d’azione cardiaco.
Potenziale d’azione cardiaco
Differenze tra muscolo striato e cardiaco
- La durata è una delle differenze più evidenti. Nelle fibre nervose si parla di 2 ms, nel muscolo striato 5-6 ms, nel miocardio, anche quando è rapido, il PdA non dura meno di 100 ms.
- Le fibre muscolari scheletriche sono organizzate in sincizi strutturali (cellule fuse tra loro in periodo prenatale in utero) a formare lunghe cellule cilindriche polinucleate. Il miocardio invece è fatto da piccole cellule mononucleate (al massimo con 2-3 nuclei).
- Il muscolo scheletrico è strutturato per generare forza in una sola direzione, mentre il miocardio genera forza in tre dimensioni. Questo fa sì che non si sviluppi forza, ma pressione, e non accorciamento quanto variazione di volume.
- Il motoneurone stimola le fibre muscolari scheletriche trasmettendo il PdA in rapporto 1:1 con meccanismo, come visto precedentemente, del tipo tutto o nulla. Ovvero un PdA arriva al motoneurone, che lo passa a tutta la fibra muscolare che si depolarizza e si contrae. Nel miocardio non ci sono sinapsi, bensì si parla di sincizio funzionale (N.B. non strutturale come per le fibre muscolari). Ciò comporta basse resistenze elettriche tra un miocita e l’altro, così che l’impulso elettrico si propaghi in modo armonico e rapido in tutto il muscolo cardiaco.
- Non sono presenti sinapsi elettriche (vedi motoneurone), bensì il PdA insorge da un centro pacemaker, cellule cardiache specializzate che generano l’input di depolarizzazione e contrazione di tutto il cuore. Il SNC non genera questi stimoli, può soltanto regolarli.
- Il muscolo cardiaco vede due tipi di PdA, uno lento (tipico delle cellule pacemaker, detto miocardio di conduzione) e uno veloce (tipico del miocardio di lavoro).
Fasi del potenziale d’azione cardiaco


Fase 0 (fase di depolarizzazione rapida)
Come avviene a livello scheletrico e neuronale, uno stimolo elettrico porta a una rapida depolarizzazione di membrana fino a un valore soglia che comporta l’apertura dei canali al sodio voltaggio-dipendenti. Questo comporta correnti entranti al sodio (molto intense, circa 500 nA, nano-ampere) che in 1-2 ms depolarizzano la membrana fino a valori di circa +20/30 mV. Questa fase è particolarmente importante nel miocardio di lavoro (PdA veloce), mentre è assente nel miocardio di conduzione (PdA lento) (figura 6). Infatti, in quest’ultimo, la depolarizzazione è soltanto calcio dipendente.
Fase 1 (fase di ripolarizzazione rapida iniziale)
I canali al sodio voltaggio dipendenti della fase 0 si chiudono a favore di correnti uscenti ripolarizzanti al potassio, dette Ito1, ovvero transient outward 1. Questi canali sono rapidissimi, si aprono e subito si richiudono. Inoltre, sono molto espressi nell’epicardio, dove la fase 1 è molto pronunciata. Sono meno espressi nell’endocardio. Inoltre, è presente un’altra corrente uscente, al cloro, detta transient outward 2 (Ito2). Il cloro ha un potenziale di equilibrio di circa -90 mV, infatti, come il potassio, è ripolarizzante.
Fase 2 (fase di plateau)
Questa fase si caratterizza per la coesistenza di 2 correnti: una al potassio detta delayed rectifier (IKs); e una al calcio detta slow inward current (ICa-L). Il calcio ha un potenziale di equilibrio positivo causando depolarizzazione. Inoltre, questi canali sono voltaggio dipendenti, ma hanno una cinetica di apertura lenta, per questo detti slow. Per cui, lo stesso impulso che fa aprire i canali al sodio della fase 0 stimola l’apertura anche questi canali, ma che si aprono più tardi. La coesistenza di una corrente depolarizzante e ripolarizzante fa si che il potenziale si mantenga per tanti millisecondi costante tra 0 e -20 mV.
Una corrente fondamentale è IK1, detta inward rectifier o anomalous rectifier. Tipicamente le correnti al potassio sono uscenti, mentre in questo caso c’è una preferenza a far entrare potassio. In realtà ciò avverrebbe soltanto se il potenziale di membrana fosse più negativo del potenziale di equilibrio del potassio (-100 mV), situazione paradossale che non si presenta mai in condizioni fisiologiche. Tra -100 mV e 0 mV il canale fa passare correnti uscenti ripolarizzanti. Tuttavia, il canale si chiude a 0 mV in un’ottica di risparmio energetico. Infatti, il PdA cardiaco dura molti millisecondi, rischiando che le correnti che si instaurano causino uno squilibrio ionico tra ambiente intra e extra cellulare, che dovrebbe essere ripristinato da pompe ATP dipendenti. Allora, per tanti canali che si aprono (sodio, potassio, cloro ecc…) altrettanti devono chiudersi. A questo ci pensano questi canali, facendo così durare più lungo possibile la fase di plateau.
Fase 3 (fase di ripolarizzazione rapida terminale)
I canali al calcio si chiudono in modo tempo-dipendente. La risultante è una corrente ripolarizzante uscente al potassio che ripolarizza la membrana.
Fase 4 (fase diastolica)
Nelle cellule a risposta rapida in questa fase si arriva al valore di riposo di membrana di circa -80/-90 mV in attesa di una nuova depolarizzazione. Una situazione diversa si riscontra nelle cellule a risposta lenta (le cellule pacemaker) dove il potenziale di riposo è instabile (figura 6), caratterizzata da una graduale depolarizzazione dovuta alla corrente pacemaker o corrente funny (If). Questa corrente è cAMP dipendente, caratteristica alla base della modulazione neuronale della frequenza cardiaca. E’ un canale misto sodio-potassio appartenente alla famiglia dei canali del potassio, che si apre in ripolarizzazione e si chiude con depolarizzazioni. A causa di questa caratteristica è detto canale autolimitante a feedback negativo. Infatti, lasciando passare correnti entranti di sodio causa depolarizzazione di membrana che fa chiudere il canale promuovendo la ripolarizzazione di membrana. Questo però lo fa riaprire e inizia il ciclo di apertura-chiusura.
Metodi di studio dei canali ionici e variazioni del potenziale di membrana
Current clamp
Questo esperimento permette di osservare come varia il potenziale di membrana in risposta al passaggio di flussi ionici. Consiste nell’applicare dei microelettrodi a una cellula eccitabile (per esempio un neurone) che siano capaci di iniettare corretti e valutare la differenza di potenziale che consegue. Come rappresentato in figura 7, il microelettrodo eI inietta corrente, mentre quello eV misura la differenza di potenziale tra interno ed esterno della cellula originata secondo la legge di Ohm, dalla relazione ∆V=i x R. Per disegno sperimentale l’esterno viene messo a terra, ovvero 0 mV. Iniettando una corrente essa assume una forma a “onda quadra”, mentre la differenza di voltaggio registrata ha un aspetto esponenziale. Ciò si deve a due fenomeni: la membrana non ha una sola resistenza, ma è influenzata da più canali e quindi più ioni con proprietà elettrochimiche diverse; inoltre, è presente una deformazione temporale.

Voltage clamp
Questo metodo, detto di blocco del voltaggio, consente di fissare il potenziale di membrana di una cellula eccitabile su valori decisi dallo sperimentatore per registrarne gli effetti in termini di correnti. Questo esperimento venne usato dai ricercatori Hodgkin e Huxley a metà del ‘900 per definire le correnti che caratterizzano il potenziale d’azione.
Utilizzando infatti un assone gigante di calamaro immerso in una soluzione salina, applicarono due fili di metallo, uno dentro all’assone e uno nella soluzione esterna, collegati per misurare il potenziale di membrana e rilevarlo con un voltimetro. A questo punto un amplificatore operazionale rileva una seconda differenza di voltaggio calcolata tra il potenziale di membrana appena rilevato e una pinza di voltaggio settata dallo sperimentatore (command voltage in figura 8). Facendo un esempio, consideriamo che il potenziale di riposo della membrana sia -70 mV. Per fissare tale valore a 0 mV, dobbiamo impostare la pinza di voltaggio a +70 mV, così che la differenza di voltaggio tra -70 mV e +70 mV sia 0 mV. Dopodiché con un misuratore di corrente si registrano le correnti che si formano.
Questo esperimento è fondamentale sia per valutare i fenomeni di potenziale d’azione passivi (bloccando il potenziale di membrana su valori sotto-soglia), sia attivi (bloccando il potenziale di membrana su valori sopra-soglia). In questo modo Hodgkin e Huxley osservarono che depolarizzando la membrana si instaurano correnti al sodio, mentre per valori di riposo predominano correnti al potassio.

Patch clamp
E’ quella tecnica usata per misurare le correnti ioniche attraverso i canali ionici. Gli elettrodi utilizzati sono micropipette con una punta in vetro molto grande e smussata che consentono, se avvicinate a una cellula, di creare un sigillo con la membrana. La punta delle micropipette è riempita di una soluzione salina contenente un filo di argento clorurato (formando così un micrelettrodo) collegato a un amplificatore elettronico. questa configurazione, detta cell-attached (o cellula adesa), permette di isolare un singolo canale ionico nella punta della micropipetta. In questo modo la soluzione nella micropipetta simula il liquido extracellulare, situazione detta inside-out. Se si applica una leggera pressione negativa con la micropipetta, si crea la configurazione whole-cell (detta outside-out) con cui è possibile simulare il liquido intracellulare nella punta della micropipetta. Inoltre, è possibile registrare le correnti provenienti da tutti i canali (non solo uno come nel caso cell-attached).

Fonti:
- Carbone Emilio, Cicirata Federico, Aicardi Giorgio; Fisiologia: dalle molecole ai sistemi integrati; EdiSES (2009).
- Anna Belfiore, Chiara Berteotti, Gerardo Biella, Mario Rosario Buffelli, Barbara Colombini, Marcello D’Ascenzo, Camillo Di Giulio, Riccardo Fesce, Gianfranco Franchi, Silvia Giovedì, Marco Luppi, Valerio Magnaghi, Sergio Masetto, Andrea Moriondo, Giuseppina Mudò, Agostino Palmeri, Daniela Puzzo, Vittorio Ricci, Mario Rosanova, Giulio Alfredo Sancini, Mariarosaria Santillo, Francesca Uberti, “Fisiologia umana – fondamenti”, edi-ermes (2018).
Crediti immagini:
- Immagine in evidenza: https://news.harvard.edu/gazette/story/2020/12/how-neurons-form-long-term-memories/
- Figura 1: Carbone Emilio, Cicirata Federico, Aicardi Giorgio; Fisiologia: dalle molecole ai sistemi integrati; EdiSES (2009).
- Figura 2: https://www.biopills.net/potenziale-di-membrana-potenziale-azione/
- Figura 3: Carbone Emilio, Cicirata Federico, Aicardi Giorgio; Fisiologia: dalle molecole ai sistemi integrati; EdiSES (2009).
- Figura 4: Anna Belfiore, Chiara Berteotti, Gerardo Biella, Mario Rosario Buffelli, Barbara Colombini, Marcello D’Ascenzo, Camillo Di Giulio, Riccardo Fesce, Gianfranco Franchi, Silvia Giovedì, Marco Luppi, Valerio Magnaghi, Sergio Masetto, Andrea Moriondo, Giuseppina Mudò, Agostino Palmeri, Daniela Puzzo, Vittorio Ricci, Mario Rosanova, Giulio Alfredo Sancini, Mariarosaria Santillo, Francesca Uberti, “Fisiologia umana – fondamenti”, edi-ermes (2018).
- Figura 5: https://app.testammissione.com/potenziale-dazione-cardiaco/
- Figura 6: https://moodle2.units.it/pluginfile.php/100090/mod_resource/content/1/7%20Lautoritmicità.pdf
- Figura 7: https://slideplayer.it/slide/524946/
- Figura 8: Carbone Emilio, Cicirata Federico, Aicardi Giorgio; Fisiologia: dalle molecole ai sistemi integrati; EdiSES (2009).
- Figura 9: Carbone Emilio, Cicirata Federico, Aicardi Giorgio; Fisiologia: dalle molecole ai sistemi integrati; EdiSES (2009).