Le leggi dell’ereditarietà spiegano come si trasmettono i caratteri da una generazione all’altra. Nel corso del ‘900 nacque la genetica classica, che si occupò principalmente di approfondire e correggere gli studi di Mendel. Infatti, i ricercatori si accorsero che spesso la realtà è più complessa di quanto osservato da Mendel. In quegli anni si individuarono alcuni caratteri che dipendevano dalle interazioni tra alleli diversi dello stesso gene o tra geni diversi. Inoltre, spesso, i caratteri erano influenzati dall’ambiente. Addirittura, si identificarono casi in cui il fenotipo era determinato da più geni e altri in cui un solo gene influenzava più caratteri. Inoltre, gli alleli non sempre mostrano un semplice rapporto di dominanza e recessività. Nonostante ciò, le leggi mendeliane restano sempre valide e si definisce “eccezione” ciò che non può essere spiegato dalle teorie di Mendel.
Eccezioni alle leggi di Mendel: le interazioni alleliche
Dominanza Incompleta
La dominanza incompleta fa parte delle eccezioni alle leggi di Mendel, in quanto l’eterozigote manifesta un fenotipo intermedio tra i due fenotipi omozigoti. Nella dominanza incompleta, non esistono alleli dominanti e alleli recessivi, contrariamente a quanto descritto da Mendel nei suoi studi. Infatti, entrambi gli alleli partecipano in egual misura a determinare il fenotipo dell’organismo eterozigote. L’eterozigote possiede una copia di ciascun allele per cui produce una quantità di trascritto e tradotto non sufficiente ad esprimere a pieno il fenotipo omozigote. Solo in omozigosi la presenza “doppia” dell’allele permette di esprimere a pieno il fenotipo previsto. Inoltre, incrociando due eterozigoti, il rapporto fenotipico nella progenie è di 1:2:1, non il classico rapporto 3:1 scoperto da Mendel nei suoi esperimenti. Difatti, il rapporto 3:1 è tipico di quei caratteri che presentano un allele che domina sull’altro, per cui il fenotipo dell’eterozigote è identico all’omozigote dominante.
Bella di notte
Mirabilis jalapa, meglio conosciuta come “bella di notte“, è una pianta originaria dell’America meridionale, in particolare del Perù. Il colore dei petali è determinato da due alleli, indicati nella Figura 1 come R e r. L’omozigote RR presenta fiori rossi, l’omozigote rr invece ha i fiori bianchi. Incrociando le due linee pure, RR e rr, si ottiene una generazione filiale (F1) di eterozigoti Rr a fiori rosa. L’eterozigote Rr presenta un fenotipo intermedio tra i due omozigoti della linea parentale. Per di più, incrociando tra loro piante eterozigoti, si ottengono piante a fiori rossi, rosa e bianchi secondo il rapporto 1:2:1.

Codominanza
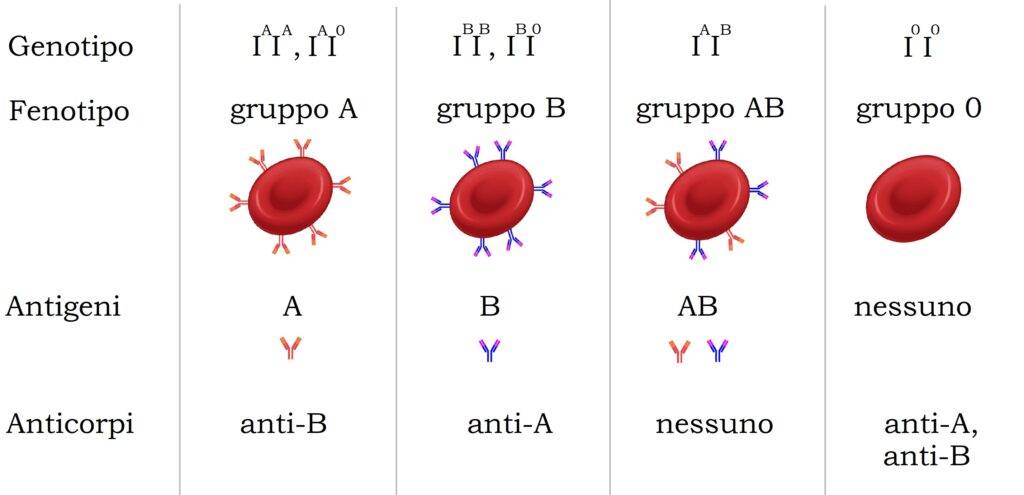
La codominanza è una delle eccezioni alle leggi di Mendel perché l’eterozigote esprime contemporaneamente entrambi i fenotipi omozigoti. Nella codominanza non esiste un allele dominante e uno recessivo. Nella codominanza si parla di allelia multipla o poliallelia, ovvero a un solo carattere fenotipico corrispondono più di due alleli dello stesso gene. Possiamo prendere come esempio i gruppi sanguigni del sistema AB0. Il gruppo sanguigno è determinato da tre alleli: IA, IB e I0. Gli alleli IA e IB sono codominanti, infatti l’eterozigote IAIB manifesta entrambi i fenotipi generando il gruppo sanguigno AB. L’allele I0, al contrario, è recessvo rispetto agli alleli IA e IB. Di fatti, l’eterozigote IAI0 sarà di gruppo sanguigno A mentre l’eterozigote IBI0 avrà il gruppo sanguigno B.
Penetranza ed espressività variabile
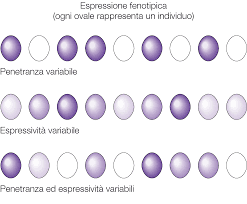
La penetranza è la frequenza con cui, dato un certo genotipo, si manifesta il fenotipo atteso all’interno di una popolazione di individui. Pertanto, per penetranza incompleta si intende quel fenomeno in cui, all’interno di una generazione di individui eterozigoti, alcuni manifestano il fenotipo recessivo. Alcune malattie genetiche sono a penetranza incompleta, ovvero la patologia non si manifesta nell’eterozigote che invece dovrebbe essere fenotipicamente malato.
Il termine espressività indica l’intensità con cui si manifesta un certo fenotipo. Per espressività variabile si intende che individui con lo stesso genotipo manifestano gradi diversi dello stesso fenotipo. Stando alle teorie di Mendel, in seguito all’incrocio tra due organismi omozigoti, si ottiene una progenie di individui eterozigoti che esprimono il fenotipo dominante. Nel caso dell’espressività variabile, alcuni ibridi ottenuti manifestano parte del fenotipo recessivo. Questo fenomeno si verifica perché l’allele dominante non è così “forte” da mascherare l’omologo. Parlando di malattie genetiche, ciò significa che gli individui eterozigoti esprimono la malattia con una gravità variabile.
Corea di Hungtington: penetranza ed espressività variabile
La corea di Huntington è una malattia rara neurodegenerativa a carico del sistema nervoso centrale. La patologia è causata da una mutazione a carico del gene HD che consiste nella ripetizione di triplette CAG lungo la sequenza di DNA. La mutazione è a carico del gene codificante per la proteina “huntingtina”, la quale risulta alterata causando la morte dei neuroni in diverse aree cerebrali. La corea di Huntington è una malattia genetica a carattere autosomico dominante con espressività e penetranza variabile. Infatti, non tutti i soggetti eterozigoti manifestano la malattia (penetranza incompleta), inoltre possono manifestare la patologia con un diverso grado di gravità (espressività variabile).
La sovradominanza come eccezione alle leggi di Mendel
Per sovradominanza o eterosi si intende il fenomeno per cui gli individui eterozigoti manifestano un fenotipo migliorato rispetto agli omozigoti. Tale condizione è nota anche come vigore dell’ibrido o vantaggio dell’eterozigote. Si tratta di uno dei tanti fenomeni genetici che esulano dalle leggi dell’ereditarietà mendeliana. Per comprendere meglio l’eterosi, basta pensare alla condizione opposta cioè la depressione da consanguineità o inbreeding depression. In genetica, per inbreeding o inincrocio si intende l’accoppiamento tra consanguinei il cui grado di parentela risalga a tre o quattro generazioni precedenti. Più il genotipo dei due individui è simile e più è probabile che la prole manifesti un fenotipo difettoso o svantaggiato a livello selettivo. Questo fenomeno è noto da secoli, infatti, già nel 1877 Darwin scrisse: “la fecondazione incrociata è generalmente positiva, l’autofecondazione invece porta spesso ad effetti deleteri“.
L’anemia mediterranea
Un esempio di sovradominanza è il morbo di Cooley, noto anche come anemia mediterranea o beta-talassemia. Il nome di questa anemia ereditaria deriva dalla sua diffusione nelle regioni che si affacciano sul Mediterraneo, in particolare Nord Africa e Medio Oriente. In Italia, la maggiore incidenza è in Sardegna. La beta-talassemia è causata dalla mutazione dei geni che regolano la produzione delle catene beta, uno dei due tipi proteici che costituiscono l’emoglobina. Si tratta di una seria patologia del sangue che comporta la distruzione precoce dei globuli rossi e carenza di emoglobina. A ciò segue una ridotta ossigenazione di tessuti, organi e muscoli.
L’eterosi nell’anemia mediterranea
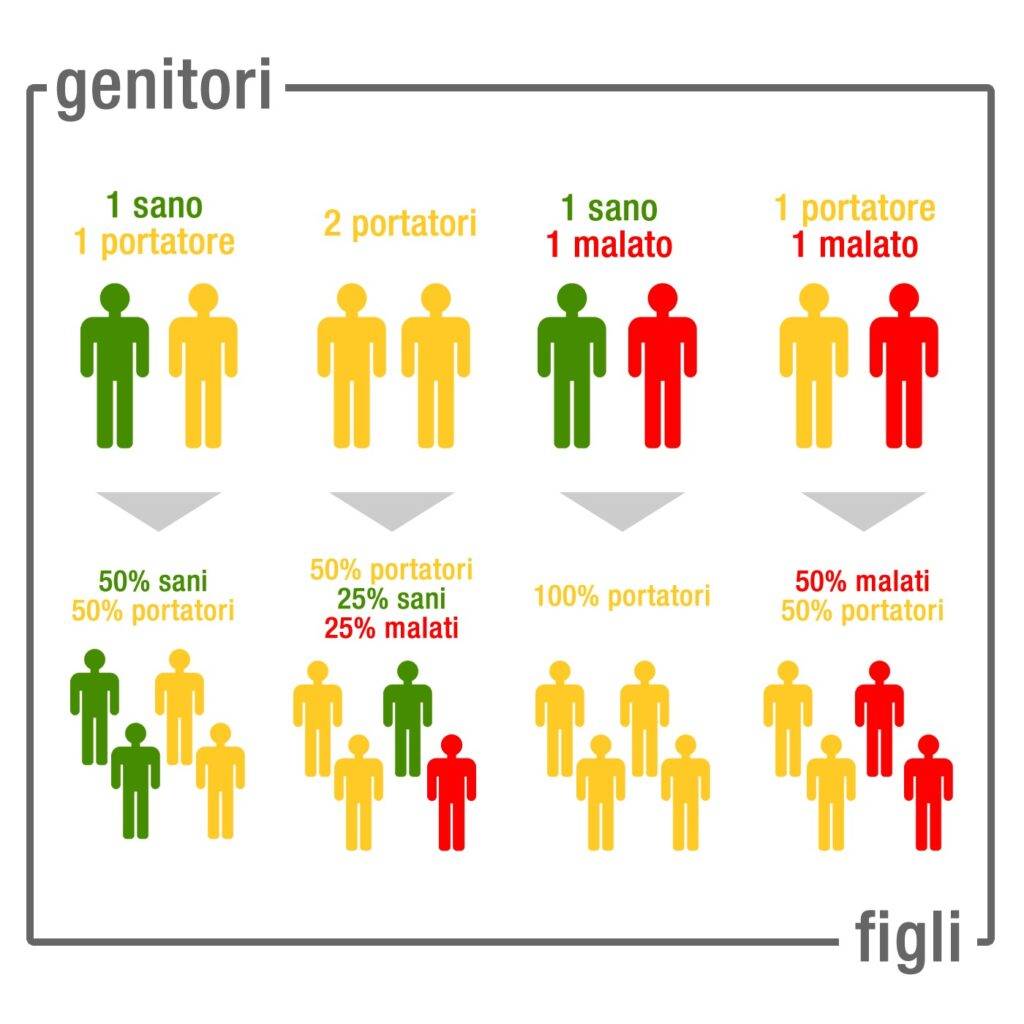
L’anemia mediterranea si manifesta in forma grave nell’individuo omozigote e prende il nome di beta-talassemia major. I sintomi compaiono a circa 6 mesi di vita e sono: debolezza, pallore, ittero, deformità delle ossa del volto, crescita lenta, gonfiore addominale, urine scure. A volte si verifica la splenomegalia, ovvero l’ingrossamento della milza. Chi soffre di beta-talassemia major deve sottoporsi a frequenti trasfusioni di sangue. D’altra parte, le ripetute trasfusioni provocano accumuli di ferro che possono causare un’intossicazione da ferro. Di conseguenza, il paziente può sviluppare siderosi miocardica e scompenso cardiaco, che possono portare al decesso del paziente. Nel caso dell’anemia mediterranea, l’eterozigote è avvantaggiato a livello fenotipico, infatti i sintomi della malattia sono lievi (beta-talassemia minor) o del tutto assenti. Pertanto, l’individuo eterozigote è definito portatore sano. Il problema della condizione di portatore sano non sta tanto nella sintomatologia, quanto nella possibilità di trasmettere la malattia ai figli.
Utilizzo commerciale dell’eterosi
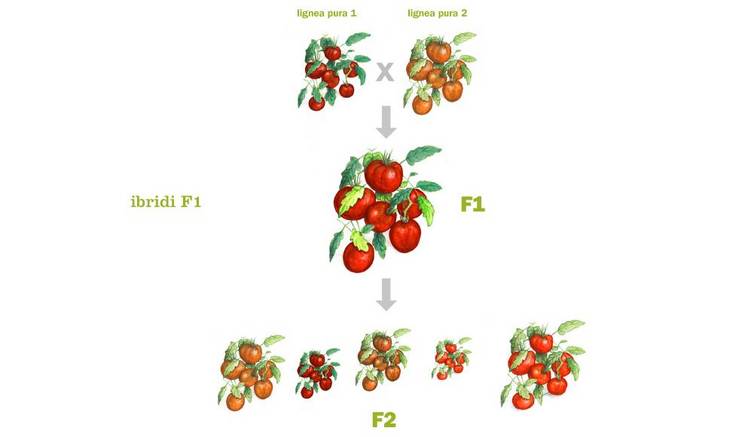
Oltre all’ambito medico, il vigore dell’ibrido è molto studiato anche in agricoltura e in zootecnia. Infatti, molti allevatori incrociano bovini da carne per ottenere degli animali più corpulenti e longevi rispetto al bestiame allevato all’interno dello stesso ceppo genetico. Anche gli agricoltori sfruttano questa tecnica per migliorare il raccolto e renderlo più redditizio. Questa pratica per rendere più redditizio il proprio allevamento o raccolto è usata da secoli. Infatti, Beal fu il primo che nel 1870 propose di coltivare mais ibrido. Già allora Beal osservò che il mais ibrido cresceva più in alto e produceva chicchi più numerosi rispetto alle linee pure generate per autofecondazione. Solamente nel 1914, con Shull, si approfondirono gli studi legati al vigore dell’ibrido nel mais. Le specie che hanno conseguito i migliori risultati nell’applicazione dell’eterosi sono le seguenti: banano, cetriolo, cipolla, cocomero, girasole, mais, melo, pomodoro, sorgo.
Eccezioni alle leggi di Mendel: le interazioni geniche
Epistasi
L’epistasi è una forma di interazione tra geni situati in un locus differente dello stesso cromosoma. Il fenomeno si verifica quando una coppia di alleli (detti epistatici) copre l’espressione fenotipica di un’altra coppia di alleli (detti ipostatici). L’individuo manifesterà un fenotipo che corrisponderà alla risultante di questa interazione genica. L’epistasi si verifica quando le due coppie di alleli dei geni sono topograficamente vicine sullo stesso paio di cromosomi. Infatti, durante la meiosi questi geni non assortiscono l’uno indipendentemente dall’altro e risulteranno associati tra loro. L’epistasi si distingue in recessiva e dominante. L’epistasi è recessiva quando è l’omozigote recessivo a mascherare l’effetto degli alleli sul gene ipostatico. Invece, l’epistasi è dominante quando l’allele dominante del gene epistatico maschera gli effetti dell’altro gene. In seguito a incrocio diibrido, per effetto epistatico, otteniamo un rapporto fenotipico di 9:3:4 invece del classico rapporto mendeliano 9:3:3:1.
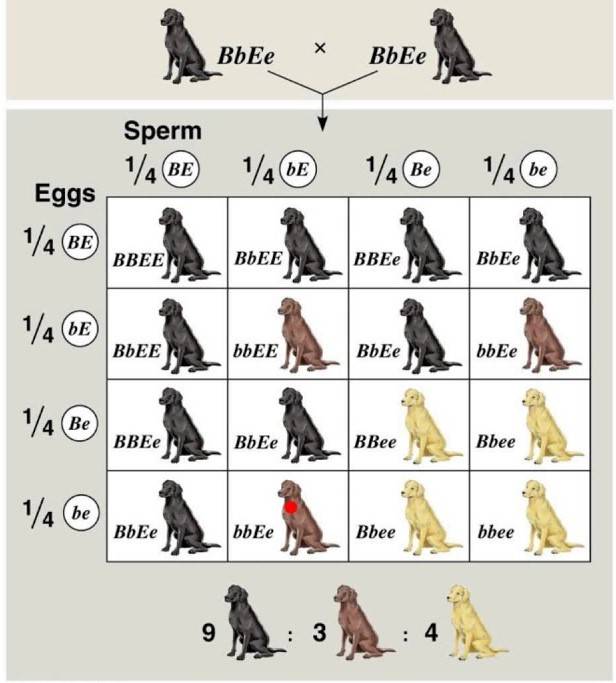
Il manto del Labrador
Il manto del Labrador può essere nero, marrone o giallo. Se i genitori sono entrambi neri, la maggior parte dei cuccioli sarà nera. Questo fenomeno è in parte descritto da Mendel. Infatti, il gene per la pigmentazione del manto è determinato da due alleli: B e b, dove B è dominante rispetto a b. Il manto marrone si manifesterà solo nell’omozigote bb, mentre i cani BB e Bb saranno neri. Nel labrador esiste anche un altro gene, posto in un locus differente rispetto al primo, che determina la deposizione o meno del pigmento. Tale gene presenta due alleli: E ed e. Se il genotipo del cane è omozigote recessivo ee, allora si inattiva il gene per il pigmento e il cane risulterà giallo. Questo è un esempio di epistasi recessiva.
Eccezioni alle leggi di Mendel: l’interazione dei geni con l’ambiente
Archibald Garrod e William Bateson scoprirono che il fenotipo di un individuo è determinato dall’interazione tra il suo genotipo e l’ambiente in cui vive. In particolare, i due scienziati furono i pionieri nella ricerca degli errori congeniti del metabolismo. Studiando alcune famiglie i cui membri presentavano anomalie nel metabolismo, essi ne identificarono la causa. Si trattava di un’alterazione genetica che impediva all’organismo di effettuare specifiche reazioni chimiche. Quando gli effetti genetici sono modulati da fattori esogeni, ad esempio lo stile di vita, esistono opportunità di prevenzione e promozione della salute. Infatti, gli scienziati osservarono come gli effetti negativi associati a tale alterazione potevano essere evitati modificando la dieta dei pazienti. In generale, l’ambiente può influenzare la trascrizione e la traduzione del DNA attraverso modifiche chimiche che regolano l’attivazione funzionale dei geni (epigenetica).
L’esempio del gatto siamese
I gatti siamesi sono gatti bianchi con aree del corpo più scure. Si tratta di una forma di albinismo parziale che deriva dalla mutazione del gene codificante la tirosinasi. La tirosinasi è l’enzima responsabile della produzione di melanina. Il gene difettoso rende la tirosinasi termolabile: non funziona alle normali temperature corporee, ma si attiva solo a basse temperature. Infatti, i cuccioli nascono quasi totalmente bianchi perché l’utero della mamma li tiene al caldo inattivando l’enzima. Invece, crescendo, il gatto siamese sviluppa una pigmentazione scura limitata ad alcune parti del corpo che sono quelle più fresche. Steve Jones, professore di genetica alla University College London, ha scritto su Lancet: “Dentro ogni siamese c’è un gatto nero che lotta per venire fuori”. Infatti, se facciamo crescere un gatto siamese in una camera fredda vedremo che il pelo diventerà scuro. Viceversa, se il siamese cresce in una serra allora diventerà un gatto bianco.

Conclusioni
Gregor Mendel è considerato il padre della genetica, in quanto fu il primo a riconoscere l’esistenza del gene: l’unità fondamentale dell’ereditarietà. Fu anche il primo ad adottare un vero e proprio metodo scientifico tramite la conduzione di esperimenti di verifica delle proprie ipotesi. Tuttavia, egli utilizzò un modello semplice su cui condurre gli esperimenti e ciò limitò i suoi studi. Infatti, i caratteri da lui analizzati si esprimono solo in due forme alternative, non sono mescolabili e non interagiscono tra loro. Le teorie di Mendel sono tutt’oggi valide e a partire da esse le conoscenze in ambito genetico si sono potute ampliare. Partendo dalle scoperte di Mendel è stato possibile identificare quelle che oggi conosciamo come eccezioni alle leggi di Mendel.
Fonti
- https://it.wikipedia.org/wiki/Dominanza_incompleta
- https://it.wikipedia.org/wiki/Codominanza
- https://mutagens.it/informati/penetranza-ed-espressivita/
- https://www.treccani.it/enciclopedia/epistasi_%28Enciclopedia-della-Scienza-e-della-Tecnica%29/#:~:text=Interazione%20tra%20geni%20non%20allelici,locus%20differente%20dello%20stesso%20cromosoma.
- https://stemeducation.nd.edu/resources/scientific-phenomena/november-phenomenon-2016
- https://www.treccani.it/enciclopedia/geni-e-ambiente-nello-sviluppo-del-comportamento_%28XXI-Secolo%29/
- https://www.thelancet.com/journals/lancet/article/PIIS0140-6736(10)61944-7/fulltext
Crediti immagini
- Immagine in evidenza: https://www.fortuneita.com/2022/02/24/biologia-della-riproduzione-servono-nuove-figure-professionali-garagna/
- Figura 1: gimp.org
- Figura 2: gimp.org
- Figura 3: https://slideplayer.it/slide/604945/
- Figura 4: https://www.thalassazione.it/trasmissione-della-talassemia/
- Figura 5: https://www.prospecierara.ch/it/animali/conoscenze/conoscenze-dettagli/news/was-sind-hybriden-und-wo-liegen-die-probleme.html
- Figura 6: https://stemeducation.nd.edu/resources/scientific-phenomena/november-phenomenon-2016
- Figura 7: https://www.tuttogreen.it/gatto-siamese-carattere-educazione-aspetto/