Caratteristiche della Membrana citoplasmatica
Ogni cellula presenta uno strato lipidico e proteico, detto membrana plasmatica (o membrana citoplasmatica, plasmalemma o anche la membrana cellulare), che definisce i confini intracellulari da quelli extracellulari e regola gli scambi molecolari con l’ambiente esterno. Questa struttura può essere chiamata anche membrana interna per distinguerla da quella esterna presente nei batteri gram-negativi. La membrana plasmatica racchiude al suo interno il citoplasma, la regione nucleare (nucleo per le cellule eucariote) e gli organuli che si trovano all’interno della cellula, necessari per le sue funzioni vitali. La sua struttura, prevalentemente lipidica e proteica, gli permette di essere resistente ma flessibile, auto-sigillante, e permeabile a molecole selezionate dalla cellula stessa. Oltre a questa forma di trasporto passivo, proteine di membrana specializzate promuovono gli scambi attraverso un trasporto che richiede un dispendio di energia.
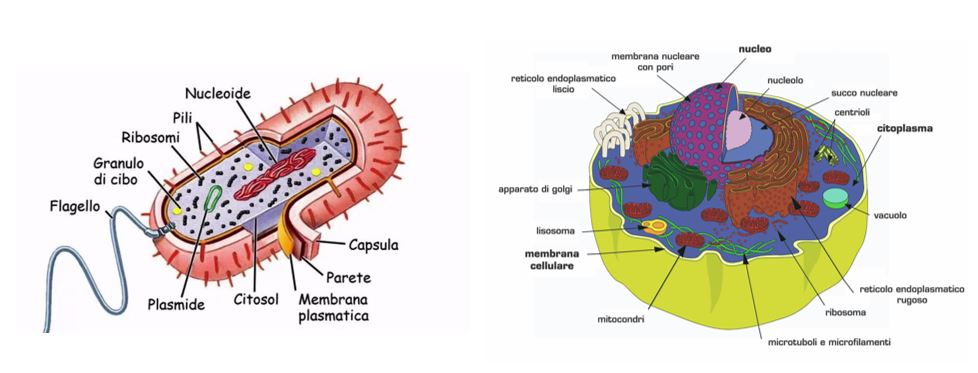
Struttura della membrana plasmatica
La membrana plasmatica è costituita (sia per i procarioti che per gli eucarioti) principalmente da due tipi di molecole: i fosfolipidi e le proteine. La differenza tra i due tipi di cellula consiste nella presenza di carboidrati e steroli nella membrana plasmatica eucariotica, elementi che le conferiscono rigidità. Non contenendo steroli, la membrana plasmatica procariotica risulta così meno rigida. L’unica eccezione si può riscontrare nei batteri del genere Mycoplasma, che presenta steroli nella membrana, ottenendo così la rigidità e compensando la mancanza di una parete cellulare.
Lo spessore della membrana può variare da 5 a 8 nm. I movimenti fluidi delle molecole fosfolipidiche e proteiche nella membrana plasmatica hanno permesso di determinare per la cellula il modello a mosaico fluido. Questa fluidità è dovuta alle interazioni non covalenti presenti tra le componenti della membrana, che consentono così alle molecole di spostarsi liberamente in senso laterale nella stessa.
Un ruolo importante in tal senso è anche determinato dal colesterolo che aumenta la flessibilità e la stabilità della membrana citoplasmatica e ne mantiene la fluidità, anche a temperature basse.
I fosfolipidi si dispongono a formare un doppio strato fosfolipidico, in cui le teste polari sono rivolte verso l’esterno della membrana, e le code non polari verso l’interno. Questa disposizione è dovuta al fatto che le code dei fosfolipidi sono code idrofobe e perciò “fuggono” dall’acqua. Le teste polari, invece, sono idrofile, e per questo rivolte verso l’esterno per interagire con la fase acquosa che si trova dentro e fuori la cellula.
Immagine della membrana cellulare
Nella seguente immagine della membrana cellulare possiamo apprezzare che la membrana cellulare è costituita da svariate e complesse proteine che ne determinano le svariate funzioni.
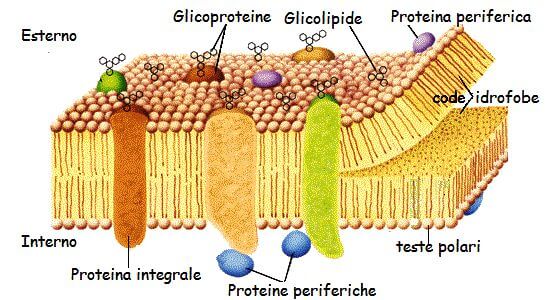
Le proteine della membrana fosfolipidica
Immerse tra i fosfolipidi si trovano le proteine di membrana, mantenute nella posizione giusta grazie a interazioni idrofobiche tra i lipidi di membrana e i domini idrofobici delle proteine. A seconda della loro disposizione all’interno della membrana, possiamo distinguere:
- Proteine periferiche, come enzimi o strutture di sostegno;
- Proteine integrali, come le proteine di trasporto.
Le proteine svolgono diverse funzioni fondamentali, come il trasporto di molecole verso l’interno o l’esterno della cellula, la funzione strutturale, la funzione enzimatica.
Fosfolipidi
I fosfolipidi di membrana sono lipidi di membrana, e come tali si prestano alla costruzione di tutte le membrane biologiche. Sono costituiti principalmente da due parti:
- Testa: costituita da glicerolo, acido fosforico e una molecola molto polare (gruppo sostituente). Il terzo carbonio C del glicerolo è legato all’acido e alla molecola polare attraverso un legame fosfodiestere: il residuo di acido fosforico esterificato al gruppo alcolico del glicerolo si lega alla molecola. Il gruppo sostituente varia a seconda della tipologia di fosfolipide, ed è una molecola polare, come colina, etanolammina ecc. Le teste dei fosfolipidi sono polari, idrofiliche (affini all’acqua).
- Code: costituite da due acidi grassi, legati attraverso un legame estere al primo e al secondo atomo di carbonio C del glicerolo. La tipologia degli acidi grassi varia a seconda del tipo di fosfolipide. Le code sono apolari, idrofobiche (fuggono dal contatto con l’acqua).
I fosfolipidi nella costruzione della membrana plasmatica e cellulare
Le differenze presenti tra teste e code determinano la disposizione dei fosfolipidi nella costruzione della membrana. Posti in una soluzione acquosa, i fosfolipidi si dispongono a formare una struttura in cui le code (in quanto idrofobiche) si posizionano verso l’interno, lontano dell’acqua. Le teste idrofiliche stanno bene a contatto con l’acqua, e si posizionano verso l’esterno a chiudere la struttura. Per questa loro caratteristica, i fosfolipidi sono considerati molecole anfipatiche.

Le strutture dei fosfolipidi
I fosfolipidi in acqua possono dare forma a strutture differenti:
- Micelle: strutture sferiche, costituite da una decina fino a qualche centinaio di molecole;
- Doppio strato: due strati di fosfolipidi costituiscono la struttura a doppio strato fosfolipidico da noi già conosciuta, con le code idrofobiche all’interno e le teste idrofiliche all’esterno. Essendo una struttura aperta, presenta i bordi esposti all’acqua e per questo motivo risulta instabile. Spontaneamente forma un aggregato lipidico detto vescicola.
- Vescicole: dette anche liposomi; queste strutture possono inglobare acqua in uno scompartimento interno, separato dalle code idrofobiche.

La biosintesi dei fosfolipidi inizia con l’acilazione con acil-coenzima A di una molecola di glicerolo-3- fosfato. Si forma acido fosfatidico, precursore di tutti i fosfolipidi e da cui deriva anche il nome.
I fosfolipidi possono essere degradati in un organismo dagli enzimi idrolitici fosfolipasi, che catalizzano la scissione dei legami estere.
Proteine di membrana
Le proteine di membrana si trovano immerse tra i fosfolipidi nel mosaico fluido. La posizione delle proteine nella membrana è mantenuta principalmente da interazioni idrofobiche instaurate tra i lipidi e i domini idrofobici delle proteine. Questa posizione può essere periferica o interessare tutto lo spessore della membrana. In base a questa disposizione, le proteine sono definite:
- Proteine periferiche: si trovano nella parte superficiale della membrana, come proteine enzimatiche o strutturali;
- Proteine integrali: si trovano immerse nella membrana e, in alcuni casi la attraversano da parte a parte (proteine transmembrana). Tra queste troviamo le proteine canale (pori) che formano un canale di passaggio che consente alle molecole di entrare o uscire dalla cellula. Sono, per esempio, le proteine di trasporto.
Nelle membrane cellulari non vanno dimenticate anche le proteine anfitropiche, che si trovano sia nel citosol che in associazione con le membrane; la loro affinità con la membrana è data dal legame non covalente con una proteina o lipide di membrana.
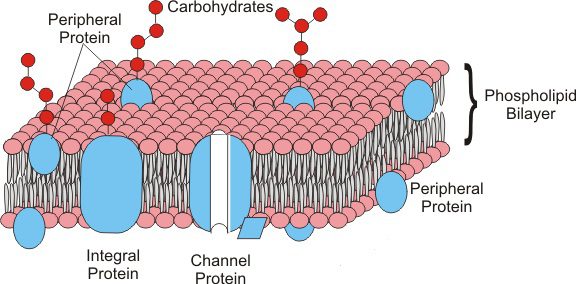
Funzione della membrana plasmatica
Le funzioni prevalentemente svolte dalle proteine di membrana sono:
- Enzimatica: catalizzano le reazioni chimiche che avvengono a livello della membrana;
- Di sostegno o strutturale: mantengono la struttura della membrana plasmatica;
- Trasporto: permette di traslocare le molecole verso l’interno o l’esterno della cellula;
- Interazioni cellulari: sono coinvolte nella comunicazione tra cellule;
- Riconoscimento: hanno strutture che consentono il riconoscimento di una molecola;
- Recettori: hanno strutture che permettono loro di ricevere e riconoscere segnali, come ormoni, in seguito ai quali possono essere avviati determinati processi cellulari.
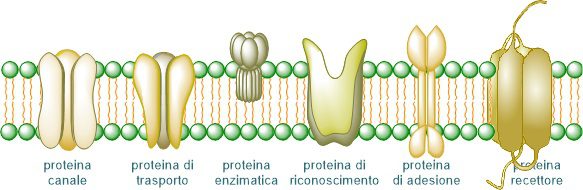
Le proteine della membrana cellulare
Le proteine di membrana possono essere associate a carboidrati (glicoproteine) o a lipidi (lipoproteine). Le glicoproteine sono abbondanti nella membrana, ed espongono i propri gruppi glucidici verso l’esterno della cellula dove, insieme ai gruppi glucidici di glicolipidi, costruiscono un rivestimento glucidico sulla superficie cellulare detto glicocalice. Grazie alla formazione di legami idrogeno, i gruppi glucidici aiutano a conferire stabilità alla membrana plasmatica. Oltre a questa funzione, le glicoproteine sono recettori di membrana per ormoni e fattori di crescita, intervengono nelle interazioni cellulari, sono siti di riconoscimento di batteri e virus.
Le lipoproteine nei batteri svolgono ruoli importanti nella fisiologia batterica, e sono ancorate perifericamente alla membrana plasmatica. Attraverso la via di secrezione generale o Sec, le proteine subiscono un processo che ne comporta la modifica sequenziale di un residuo di cisteina e la scissione di un peptide di segnale mediante peptidasi. Queste lipoproteine modificate possono essere ancorate alla membrana plasmatica. I ruoli fisiologici svolti dalle lipoproteine nei batteri sono:
- Garantire la stabilità della membrana;
- Ruolo nella divisione cellulare, sporulazione, coniugazione;
- Acquisizione di nutrienti;
- Trasduzione del segnale;
- Trasporto;
- Ripiegamento delle proteine;
- Meccanismi di patogenicità: ruolo in adesione, colonizzazione, invasione, persistenza.

Funzioni della membrana plasmatica
Tra le funzioni della membrana plasmatica, una delle principali è quella di contenimento. Infatti, la membrana racchiude al suo interno tutto il contenuto cellulare, quali il citoplasma e tutti gli organi cellulari.
Le funzioni della membrana sono svolte anche dalle proteine in essa presenti; tra queste, ricordiamo:
- Interazioni extra-cellulari: comunicazione con le cellule adiacenti per mezzo di specifiche proteine;
- Riconoscimento di molecole attraverso proteine in grado di riconoscerne la struttura;
- Ricevere segnali, come ormoni utili per determinare l’avvio di processi cellulari, per esempio per innescare i fenomeni di difesa cellulare;
- Trasporto di sostanze.
Il traporto attraverso la membrana cellulare
Il trasporto attraverso la membrana può essere attivo e passivo. Prendiamo in considerazione una cellula batterica posta in una soluzione ipertonica, ovvero un ambiente in cui la concentrazione di soluti è più alta rispetto a quella presente nella cellula. La cellula subisce una riduzione del volume e si raggrinzisce perché l’acqua si muove verso l’ambiente con maggior concentrazione di soluti (osmosi). Se poniamo la cellula in una soluzione ipotonica (un ambiente in cui la concentrazione di soluti è inferiore rispetto a quella nella cellula), l’acqua andrà nella cellula, attirata dalla concentrazione superiore.
Quando l’osmosi non lo consente, per traslocare le molecole la cellula dovrà attivare il trasporto attivo mediante le proteine di membrana e dispendio di molecole energetiche (ATP).
Un’altra forma di trasporto utilizzabile in questi casi è il trasporto di gruppo, presente solo nelle cellule procariote. Permette di accumulare sostanze anche contro gradiente di concentrazione. Questa forma di trasporto richiede dispendio di energia, e comporta la modifica della sostanza durante il passaggio della membrana. Quest’ultima diventa così impermeabile alla nuova sostanza, impedendone l’uscita.
Particolari vie di traslocazione attraverso la membrana plasmatica
Nei batteri esistono due vie di traslocazione attraverso la membrana plasmatica:
- Un meccanismo accoppiato traslazionalmente, presente in tutti gli organismi e mediato da particelle di riconoscimento dei segnali;
- Meccanismo specifico per i batteri mediato dal complesso Sec e ATP.
Il complesso Sec
Sec è un complesso proteico presente in batteri come Escherichia coli, e la sua componente principale è il canale proteico di conduzione presente nella membrana plasmatica. Nei batteri questo canale è costituito dalle proteine denominate SecY, SecE e SecG. Non si conosce ancora la motivazione di due vie di traslocazione, ma si pensa sia dovuto alla presenza di un numero limitato di canali SecYEG. Secondo gli studi, i canali presenti non sarebbero sufficienti a supportare la traslocazione solamente con il meccanismo accoppiato traslazionalmente.
SecY forma nella membrana plasmatica un canale a forma di clessidra. Ai lati del restringimento centrale della clessidra ci sono dei residui alifatici a catena lunga. Durante la traslocazione, le proteine del substrato attraversano il restringimento. In stato di riposo, invece, il canale viene chiuso con un dominio plug alpha elicoidale. SecE e SecG hanno il compito di stabilizzare SecY. SecE è legato a SecY all’esterno su entrambi i lati e in sua assenza va incontro a rapida degradazione.
I canali SecYEG
SecYEG può trovarsi in tre forme:
- Chiuso: non avviene traslocazione;
- Parzialmente aperto: Il legame SecAEG e SecYEG in presenza di ADP favorisce una lieve apertura del canale;
- Aperto: il legame SecAEG e SecYEG in presenza di ATP provoca una grande dilatazione del restringimento presente nel canale.
Oltre a queste possiamo trovare SecA, che svolge la funzione di traslocazione della maggior parte di proteine di E. coli. SecA è costituito da due domini: NBD1 e NBD2. La struttura primaria di NBD1 è interrotta da una reticolazione polipeptidica, che si occupa di rintracciare il polipeptide del substrato che deve entrare nella cellula. NBD2 è composto dal dominio dell’impalcatura alpha elicoidale, che intercetta la proteina del substrato, e da quello dell’ala elicoidale alpha.
La traslocazione mediante SecA
La traslocazione mediante SecA avviene in 2 fasi:
- le proteine del substrato che devono essere traslocate vengono indirizzate nel complesso Sec;
- Traslocazione mediante SecYEG.
Attraverso l’interazione con la proteina del substrato o con SecYEG, SecA subisce un cambiamento nella sua conformazione: la reticolazione polipeptidica viene ruotata e traslata in modo tale da avvicinarsi a NBD2 e assumere una conformazione aperta. Si forma quindi una cavità tra NBD1, NBD2 e la reticolazione polipeptidica, in cui SecA si lega al substrato per la traslocazione.
Vuoi saperne di più? Leggi cos’è la membrana plasmatica anche nella versione in inglese dell’articolo.
Fonti
- Immagine: https://www.chimica-online.it/biologia/membrana-cellulare.htm
- http://www.treccani.it/enciclopedia/fosfolipidi/
- Immagine: http://www.editricesanmarco.it/articoli/sfogliabili/101-4/files/assets/seo/page16.html
- Lehninger, Nelson, Cox, I principi di biochimica di Lehninger, edizione 2011, Zanichelli Editore.
- Immagine: https://www.sutori.com/story/la-cellula-procariote–qSDvEmz9GFuWYmsyS4egmiho
- https://www.sciencedirect.com/science/article/pii/S0167488914001402
- Immagine: https://commons.wikimedia.org/wiki/File:OSC_Microbio_07_03_micelle.jpg
- https://books.google.it/books?hl=it&lr=&id=g4i-LFx2oMIC&oi=fnd&pg=PR13&dq=flagelli+e+fimbrie&ots=viHp0FV5Y_&sig=FUMfPH_VwSmFJa2u3h9ZFJJVatI&redir_esc=y#v=onepage&q=flagelli%20e%20fimbrie&f=false
- Immagine: https://www.tes.com/lessons/QToW1v9X0G4Fmg/la-cellula
- http://testdimedicina.altervista.org/blog/componente-proteica-membrana-cellulare/
- https://www.gmpe.it/biologia/membrana-plasmatica
- https://www.medicinapertutti.it/argomento/glicoproteine-di-membrana/
- membrana cellulare struttura (articolo) – wikipedia
impressionante!