Introduzione
Con il termine “fotosintesi” si identifica un processo biochimico che si svolge nelle piante verdi e in altri organismi (batteri e fitoplancton), attraverso il quale, in presenza della luce solare, vengono sintetizzate specifiche sostanze, soprattutto gli zuccheri. Si tratta di un processo anabolico, ovvero basato sull’utilizzo di molecole più piccole per formare molecole più grandi (l’opposto è il catabolismo), che parte da due precursori, l’anidride carbonica atmosferica e l’acqua; in aggiunta a ciò, permette alle piante di ottenere il nutrimento necessario per crescere, ed è quindi vitale per la loro salute e sopravvivenza.
Nelle piante le varie attività biosintetiche avvengono all’interno dei plastidi, ossia organelli presenti soltanto nelle cellule vegetali, caratterizzati da una doppia membrana, dalla capacità di riprodursi autonomamente e dalla presenza di un piccolo genoma codificante per alcune proteine.
Esistono tre gruppi di plastidi, ossia gli amiloplasti, in cui è immagazzinato l’amido (un polisaccaride che funge da riserva energetica), i cromoplasti, che sintetizzano e accumulano pigmenti come carotenoidi (pigmenti di natura lipidica, precursori della vitamina A), antocianine (coloranti appartenenti alla famiglia dei flavonoidi) e xantofille (sostanze naturali incluse nella famiglia dei carotenoidi), e infine i cloroplasti. Questi ultimi raffigurano i protagonisti di questo articolo, dal momento che sono quelli in cui si compie il processo fotosintetico; i cloroplasti contengono una serie di dischi appiattiti chiamati tilacoidi (lamelle), impilati l’uno sull’altro a formare i grana (granum al singolare), sono immersi in una sostanza acquosa con un alto contenuto di proteine (specialmente enzimi), lo stroma, e si connettono tra di loro mediante delle porzioni di membrana dette lamelle stromali (Fig. 1).
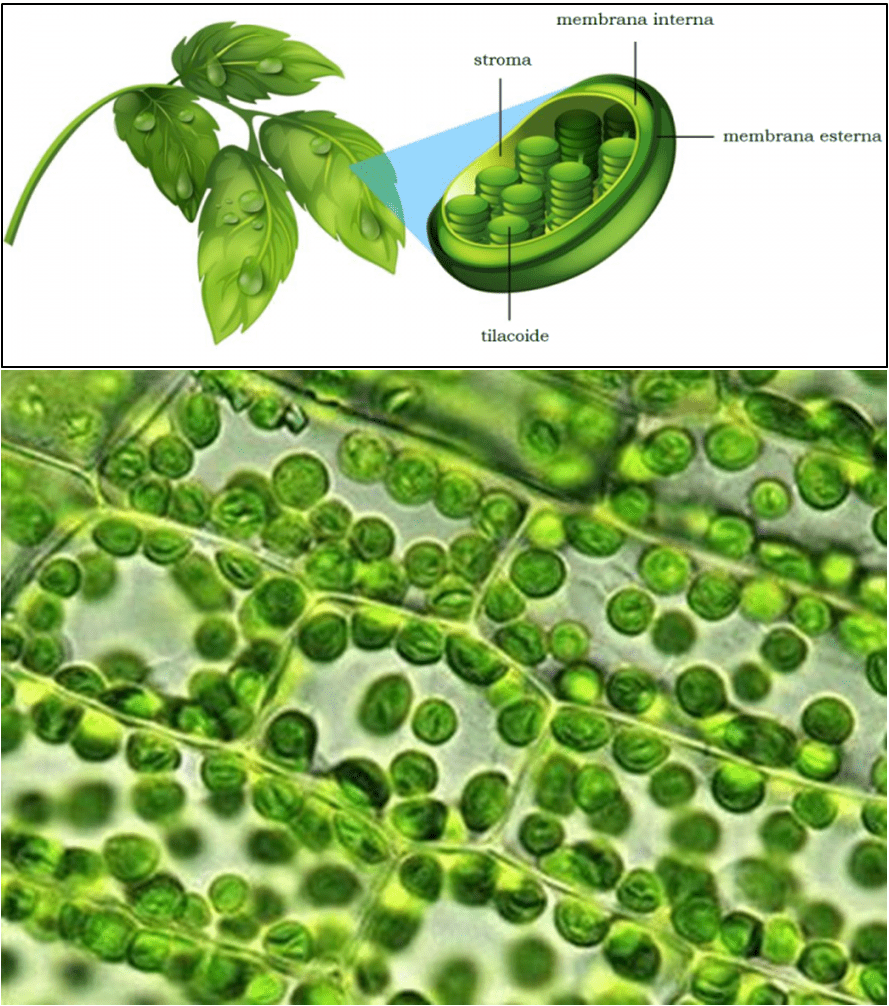
[Fonte: antropocene.it / chimica-online.it]
All’interno dei tilacoidi si trova la clorofilla, il cui nome deriva dal greco “chloros”, verde, e “phyllon”, foglia, che è la base e il motore dell’intero processo. È un pigmento verde isolato nel 1817 dai due chimici francesi Joseph Bienaimé Caventou e Pierre-Joseph Pelletier, avente una struttura eterociclica formata da un anello porfirinico con quattro anelli composti a loro volta da quattro atomi di carbonio e uno di azoto, con al centro uno ione magnesio, che mantiene rigido lo scheletro chimico al fine di impedire la dispersione dell’energia solare sotto forma di calore; legata all’anello c’è una catena idrocarburica (Fig. 2).
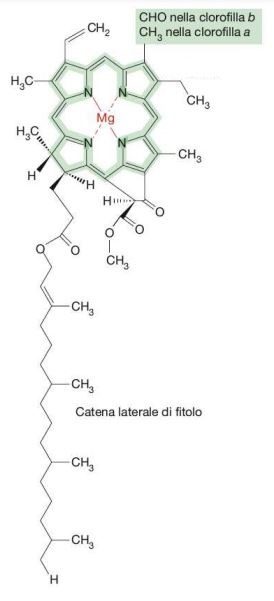
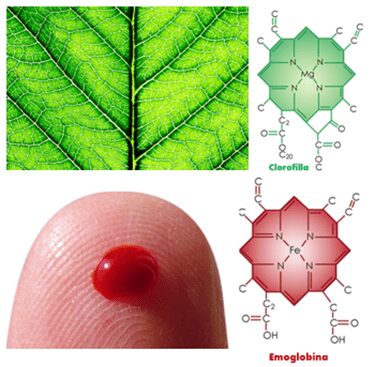
La disposizione degli elementi della clorofilla è notevolmente simile a quella del gruppo eme dell’emoglobina, con la differenza che quest’ultima contiene uno ione ferro al centro; la via biosintetica delle due molecole è la stessa (Fig. 3).
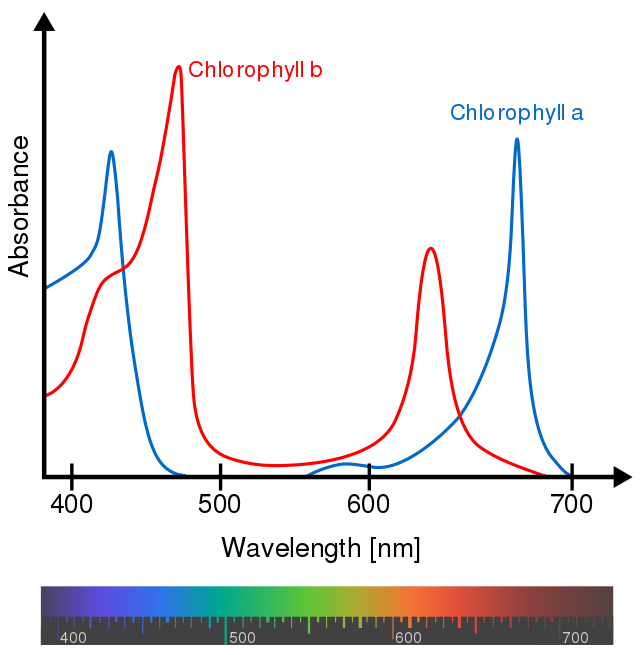
Negli eucarioti ci sono due tipi di clorofilla, ovvero clorofilla A, che assorbe specialmente la luce blu-violetta e rossa, e la clorofilla B, che cattura in prevalenza la luce blu e arancione. La ragione per cui le piante sono verdi è perché la clorofilla assorbe molto poco la lunghezza d’onda del verde (Fig. 4).
Fasi della fotosintesi
Il processo fotosintetico avviene in due fasi, ovvero la fase luminosa, che è dipendente dalla luce solare, e la fase oscura (o fase di fissazione del carbonio), che si compie in maniera indipendente dalla luce.
Fase luminosa
Iniziando dalla fase luminosa, bisogna sottolineare che nei tilacoidi la clorofilla è attorniata da complessi proteici chiamati fotosistemi, ognuno dei quali contiene da 250 a 400 molecole di pigmenti che includono, oltre alla clorofilla, i pigmenti antenna e i pigmenti accessori (carotenoidi e ficobiline). I primi assorbono la luce solare e trasmettono l’energia, per risonanza, alle molecole di clorofilla o di pigmenti accessori limitrofe, mentre i secondi assorbono i fotoni che non vengono captati dalla clorofilla e trasferiscono l’energia a quest’ultima. L’energia trasportata arriva poi nel centro di reazione (Fig. 5).
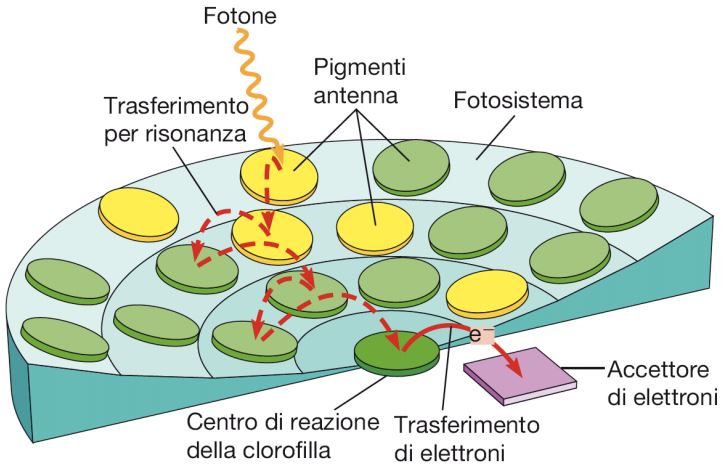
È importante evidenziare la presenza di due tipi di fotosistemi che si distinguono in base alla lunghezza d’onda della luce (indicata in nanometri) a cui gli accettori principali hanno il picco di assorbimento, e sono il fotosistema I e il fotosistema II, che assorbono rispettivamente a 700 e 680 nm (per tale ragione sono chiamati anche P700 e P680). Il centro di reazione impiega l’energia assorbita e trasferita nel fotosistema II allo scopo di indurre una separazione di cariche elettriche, una reazione di ossidoriduzione in cui la clorofilla funge da donatore di un elettrone; questo viene ceduto a un accettore primario di elettroni che si trova a un livello energetico più elevato. Gli elettroni sono generati dalla fotolisi dell’acqua, la cui reazione è descritta in questo modo:
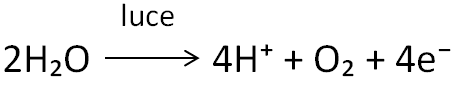
L’acqua viene scissa da un macchinario molecolare associato al fotosistema II, il complesso che libera ossigeno (o complesso che scinde l’acqua), portando alla liberazione di quattro protoni (H⁺), una molecola di ossigeno (O₂) e quattro elettroni (e⁻). A questo punto il macchinario cede gli elettroni al P680 (non tutti insieme, bensì uno alla volta), e successivamente questi vengono dislocati lungo una sequenza di molecole intermediarie che rappresentano i componenti della catena di trasporto degli elettroni.
Tali costituenti sono la feofitina (una clorofilla priva dello ione magnesio), il plastochinone A, il plastochinone B, il complesso citocromo bf e la plastocianina (proteina contenente rame). Essi ricoprono la funzione di molecole trasportatrici che trasferiscono gli elettroni a un livello energetico più basso, ovvero la clorofilla del fotosistema I. Inoltre, rilasciano energia che viene usata al fine di pompare i protoni dall’ambiente interno dei tilacoidi allo stroma creando un gradiente di concentrazione.
A fornire un canale di passaggio per i protoni è la proteina di membrana ATP-sintetasi (designata complesso CF₀CF₁, è assai somigliante al complesso F₀F₁ presente nei mitocondri), la quale utilizza l’energia potenziale del gradiente elettrochimico per sintetizzare l’ATP (adenosina 5’-trifosfato), un nucleotide che funziona da trasportatore di energia nei processi metabolici come la glicolisi (Fig. 6). La produzione dell’ATP a partire dal gradiente protonico è chiamata fotofosforilazione, e tale nome permette di distinguerla dalla fosforilazione ossidativa che avviene nei mitocondri.
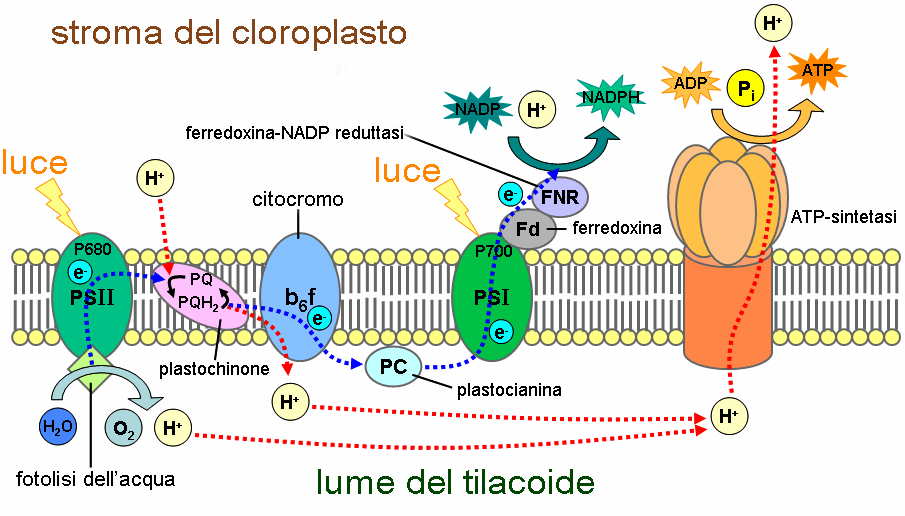
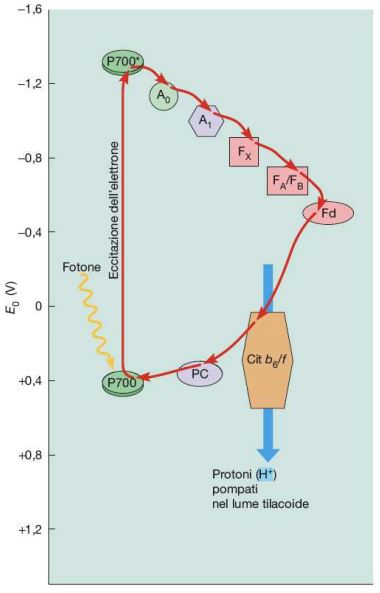
Attraverso la plastocinina, gli elettroni giungono al fotosistema I, da qui scendono lungo un’ulteriore catena di trasporto formata da altre molecole trasportatrici (il fillochinone A0, il fillochinone A1, una proteina ferro-zolfo, la ferredossina e la ferredossina NAD+ ossidoriduttasi), e in seguito passano al NADP⁺ (nicotinammide adenina dinucleotide fosfato in forma ossidata), un nucleotide che funge da cofattore per le reazioni biosintetiche e quelle di ossidoriduzione. Nel momento in cui due elettroni e un protone (H⁺) si uniscono al NADP⁺, si verifica la genesi del NADPH (in forma ridotta) (Fig. 7). Gli elettroni eliminati dal fotosistema I vengono sostituiti da quelli del fotosistema II. L’ATP e il NADPH raffigurano l’esatto prodotto della fase luminosa (2 molecole ciascuno).
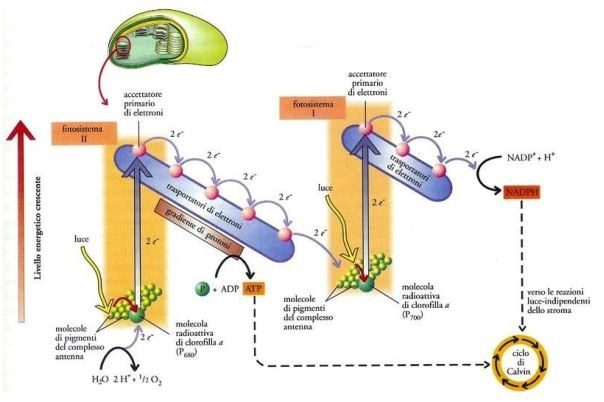
È opportuno mettere in risalto il fatto che l’ATP e il NADPH sono implicate in molteplici vie metaboliche, di conseguenza è alquanto difficile che una cellula fotosintetica richieda sempre questi due composti negli esatti rapporti realizzati dal flusso non ciclico degli elettroni. Normalmente le cellule necessitano di una mole di ATP superiore rispetto al NADPH, in quanto una buona quantità di attività cellulari funziona con l’ATP e non con il NADPH, come ad esempio il trasporto attraverso le membrane.
Ci sono circostanze in cui il consumo di NADPH è basso e/o in cui occorre una maggiore quota di ATP; per fronteggiare tale esigenza, le piante ricorrono a una via alternativa chiamata flusso ciclico degli elettroni, in cui la ferredossina dona elettroni al complesso del citocromo bf. In questo modo agli elettroni ritornano al P700 attraverso la plastocianina. Il flusso ciclico degli elettroni è associato al pompaggio unidirezionale dei protoni dal lume del tilacoide allo stroma, consentendo la sintesi di un numero più elevato di molecole di ATP (Fig. 8).
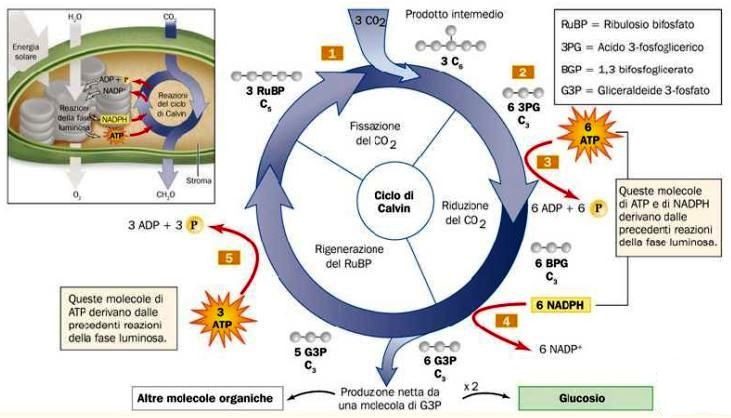
Proseguiamo adesso con la fase oscura della fotosintesi, contraddistinta dalla trasformazione dell’anidride carbonica in carboidrati (organicazione della CO₂), la cui reazione impiega l’ATP e il NADPH prodotti durante la fase luminosa. Tale fase si compie passando per tre gradini, ossia la fissazione della CO₂ nel 3-fosfoglicerato, la conversione del 3-fosfoglicerato in gliceraldeide-3-fosfato e la rigenerazione del ribulosio 1,5-bisfosfato. Questi tre stadi compongono il cosiddetto ciclo di Calvin (Fig. 9), dal nome del biochimico statunitense Melvin Calvin, vincitore del premio Nobel per la chimica grazie alle sue scoperte sulla fotosintesi.
Fase oscura
Descriviamo in dettaglio i tre step:
- Fissazione della CO₂ nel 3-fosfoglicerato: in questo primo step gioca un ruolo importante l’enzima ribulosio 1,5-bisfosfato carbossilasi (rubisco), che è collocato nello stroma dei cloroplasti (costituisce il 15% del quantitativo proteico) e catalizza il legame covalente dell’anidride carbonica al composto intermedio a cinque atomi di carbonio, il ribulosio 1,5-bisfosfato, che viene diviso in due molecole di 3-fosfoglicerato. L’enzima rubisco si trova in forma inattiva se non è carbamilato (cioè non legato a un carbammato, che è un composto avente un gruppo amminico legato a un gruppo estereo). Il processo di carbamilazione è inibito dal ribulosio 1,5-bisfosfato, che si attacca al sito attivo dell’enzima; questo, per essere messo in azione, necessita dell’intervento di un altro enzima, la rubisco attivasi, che permette la rimozione del ribulosio 1,5-bisfosfato con conseguente carbamilazione non enzimatica da parte della CO₂. In seguito avviene il legame del magnesio che innesca l’enzima rubisco.
- Conversione del 3-fosfoglicerato in gliceraldeide-3-fosfato: il 3-fosfoglicerato, generato nella prima fase, viene trasformato in un composto a tre atomi di carbonio, la gliceraldeide 3-fosfato. Tale operazione richiede la presenza di due enzimi, la 3-fosfoglicerato chinasi, che catalizza la dislocazione di un gruppo fosfato dall’ATP al 3-fosfoglicerato portando alla creazione dell’1,3-bisfosfoglicerato, e la gliceraldeide 3-fosfato deidrogenasi, che sposta due elettroni dal NADPH (che si ossida) all’1,3-bisfosfoglicerato, il quale si riduce e dà luogo a 2 molecole di gliceraldeide 3-fosfato. Una di queste viene convertita in diidrossiacetone fosfato dall’enzima trioso fosfato isomerasi.
- Rigenerazione del ribulosio 1,5-bisfosfato: nel terzo e ultimo stadio i due composti a tre atomi di carbonio si fondono e danno origine al fruttosio 1,6-bisfosfato grazie all’azione dell’enzima aldolasi. Dopodiché l’enzima fruttosio 1,6-bisfosfatasi scinde il fruttosio 1,6-bisfosfato in fruttosio 6-fosfato, con rilascio di un fosfato inorganico. Successivamente l’enzima transchetolasi catalizza lo spostamento di un gruppo chetonico dal fruttosio 6-fosfato alla gliceraldeide 3-fosfato generando due composti a cinque e a quattro atomi di carbonio, lo xilulosio 5-fosfato e l’eritrosio 4-fosfato. Quest’ultimo, grazie alla cooperazione dell’aldolasi, si unisce al diidrossiacetone fosfato con formazione di un composto a sette atomi di carbonio, il sedoeptulosio 1,7-bisofosfato. A questo punto la transchetolasi, a partire dal sedoeptulosio 1,7-bisofosfato e dalla gliceraldeide 3-fosfato, produce due pentosi fosfato (ribosio 5-fosfato e xilulosio 5-fosfato), che sono poi convertiti in ribulosio 5-fosfato, il quale diventa ribulosio 1,5-bisfosfato in seguito all’aggiunta di un gruppo fosfato da parte dell’enzima ribulosio 5-fosfato chinasi.
Nell’insieme, le molecole e le quantità di esse consumate nel ciclo di Calvin sono le seguenti: sei molecole di CO₂, sei di acqua, 18 di ATP e 12 di NADPH per generare due molecole di gliceraldeide 3-fosfato, le quali raffigurano la fonte per la sintesi del glucosio tramite un procedimento inverso alla glicolisi. Questo zucchero è indispensabile per produrre le molecole energetiche e rilasciare l’energia che permette alle piante di compiere i processi metabolici. Non bisogna dimenticare che durante la fotosintesi viene espulso come scarto l’ossigeno che, come ben sappiamo, è essenziale per la vita di tutti gli esseri viventi sulla Terra. Possiamo esprimere la liberazione di glucosio e ossigeno mediante la reazione:
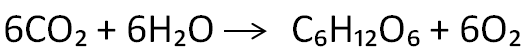
Sei molecole di acqua e sei molecole di anidride carbonica sono assorbite dalle piante e vengono trasformate in una molecola di glucosio e sei molecole di ossigeno. L’ossigeno dell’acqua si ossida, mentre il carbonio della CO₂ si riduce.
Dal momento che il ciclo di Calvin inizia con la produzione di un composto organico a tre atomi di carbonio (gliceraldeide 3-fosfato), tale processo viene detto anche via del C₃; le piante che utilizzano questo meccanismo sono chiamate piante C₃, che di solito si trovano nelle regioni temperate.
Piante C₄ e CAM
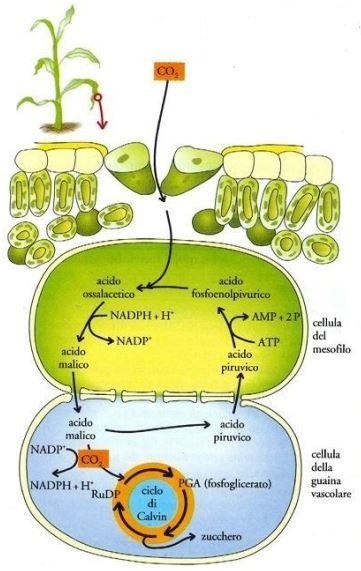
È importante adesso focalizzare l’attenzione su due categorie di piante che si distinguono in base all’attività di fissazione della CO₂, ovvero le piante C₄ e le piante CAM (metabolismo dell’acido crassulaceo). Le prime, che includono il granturco, la canna da zucchero e il sorgo, impiegano il metabolismo C₄ (o via del C₄) (Fig. 10), che si svolge nelle cellule del mesofillo (insieme dei tessuti che compongono la foglia) e nelle cellule della guaina vascolare, e inizia con il legame dell’anidride carbonica a un composto organico chiamato acido fosfoenolpiruvico con formazione di un composto a quattro atomi di carbonio, l’ossalacetato (che è implicato anche nel ciclo di Krebs).
Questa reazione è catalizzata dall’enzima acido fosfoenolpiruvico (PEP) carbossilasi. Successivamente l’enzima malato deidrogenasi ossida il NADPH a NADP⁺ e riduce l’ossalacetato a malato; questo entra nelle cellule della guaina vascolare, dove l’enzima malico lo demolisce in piruvato e anidride carbonica. La molecola di CO₂ rimossa, grazie alla rubisco, si fonde con il ribulosio 1,5-bisfosfato generando il 3-fosfoglicerato; si innesca quindi il ciclo di Calvin. Attraverso i plasmodesmi (canali di connessione tra le cellule vegetali) il piruvato ritorna nelle cellule mesofilliche e viene convertito in fosfoenolpiruvato dall’enzima piruvato fosfato dichinasi, che prende un gruppo fosfato dall’ATP, e rilascia AMP (adenina monofosfato ciclico) e due molecole di fosfato inorganico.
Nella seconda categoria di piante (CAM), che vivono in ambiente caldi e aridi, come le Crassulacee, le Cactacee (cactus), l’ananas e l’agave, si verifica un sistema diverso di fissazione fotosintetica della CO₂ (Fig. 11): nel corso della notte, quando in genere la temperatura è più bassa e l’umidità è più alta, gli stomi (strutture composte da due cellule presenti nelle foglie, nei fiori e nei fusti) delle cellule del mesofillo si aprono, consentendo la penetrazione dell’anidride carbonica. Questa si unisce al fosfoenolpiruvato conducendo alla sintesi dell’ossalacetato, che viene ridotto a malato e conservato nei vacuoli (organuli che raccolgono acqua, sostanze nutritive e sostanze di scarto). Durante il giorno gli stomi si chiudono, il malato subisce la dissociazione in piruvato e anidride carbonica, che viene fissata nel 3-fosfoglicerato ed entra nel ciclo di Calvin. Le reazioni di questa via metabolica sono catalizzate dagli stessi enzimi della via C₄.
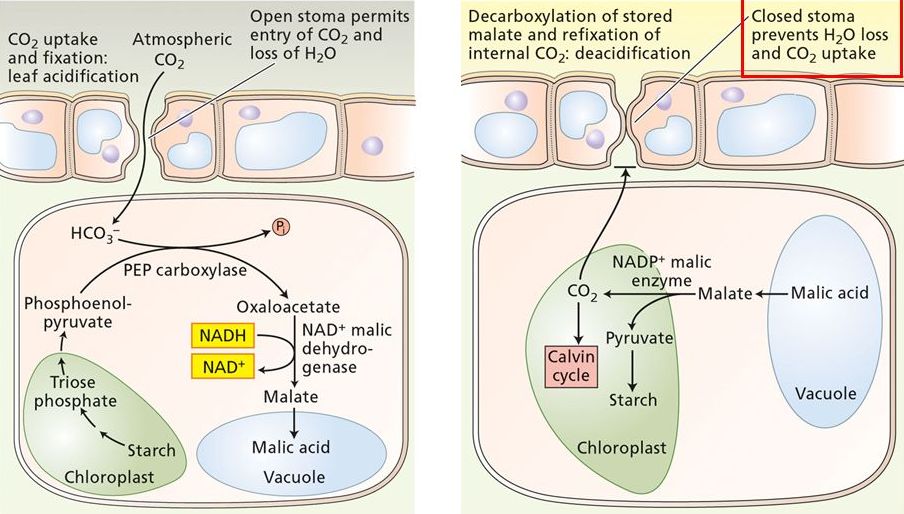
Quando il clima diventa più umido, per esempio dopo un temporale, le piante CAM impiegano il meccanismo delle piante C₃.
Fotosintesi nei batteri
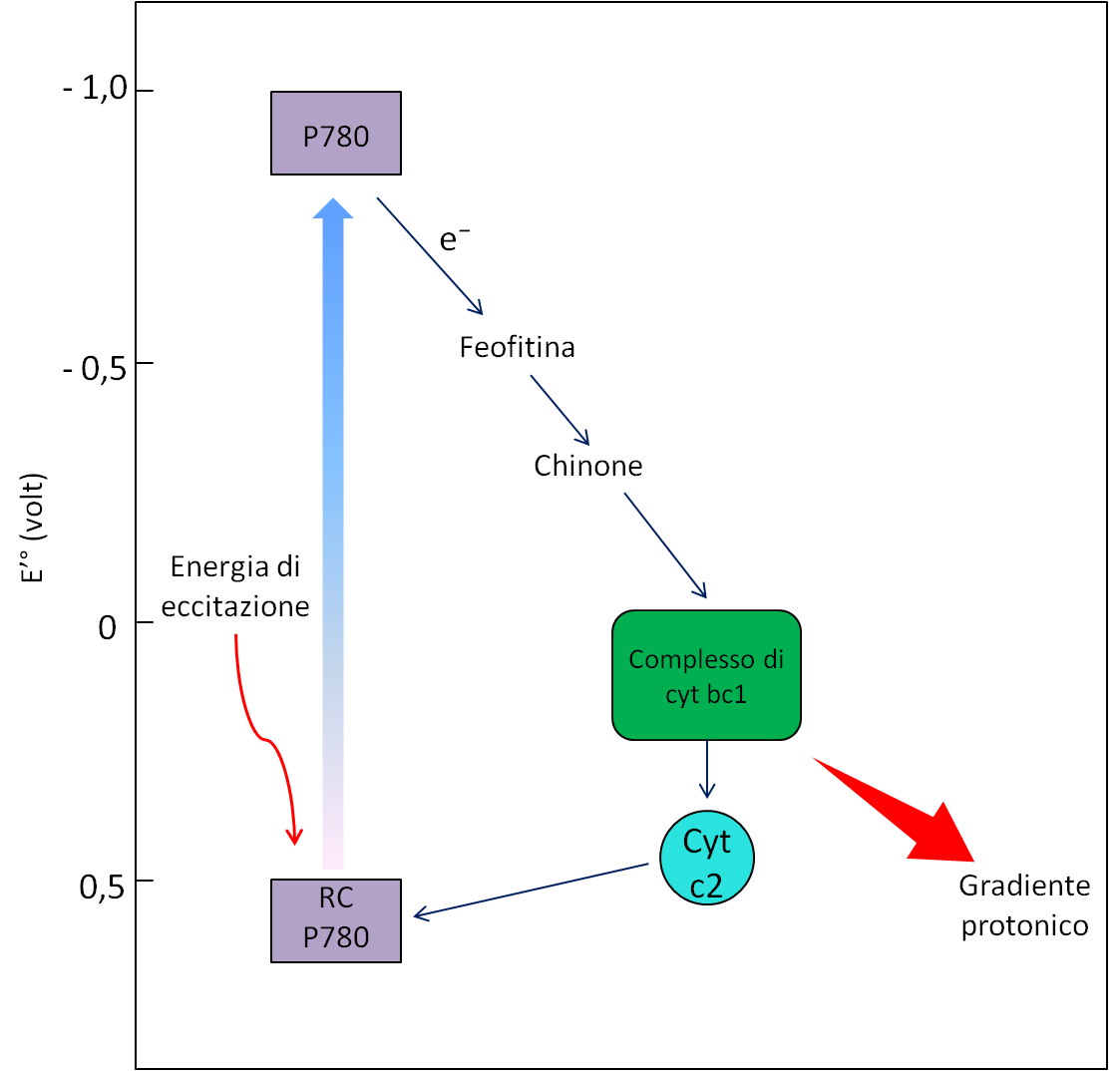
In questa parte dell’articolo esponiamo il meccanismo fotosintetico dei batteri. Alcuni di questi microorganismi possiedono un sistema di fototrasduzione abbastanza semplice in cui è presente un solo centro di reazione. I batteri su cui ci concentreremo sono i rodobatteri e i solfobatteri verdi; nel primo gruppo gli elettroni effettuano un flusso ciclico a partire dal centro di reazione P780, passano alla feofitina, a un chinone, al complesso di citocromi bc1 e, attraverso il citocromo c2, tornano al centro di reazione (Fig. 12). Nel secondo gruppo, invece, dopo l’eccitazione del centro di reazione P480, gli elettroni possono seguire due circuiti differenti:
- Un percorso ciclico nel quale vengono trasferiti a un chinone, al complesso di citocromo bc1 per poi tornare al centro di reazione grazie al citocromo c (Fig. 13);
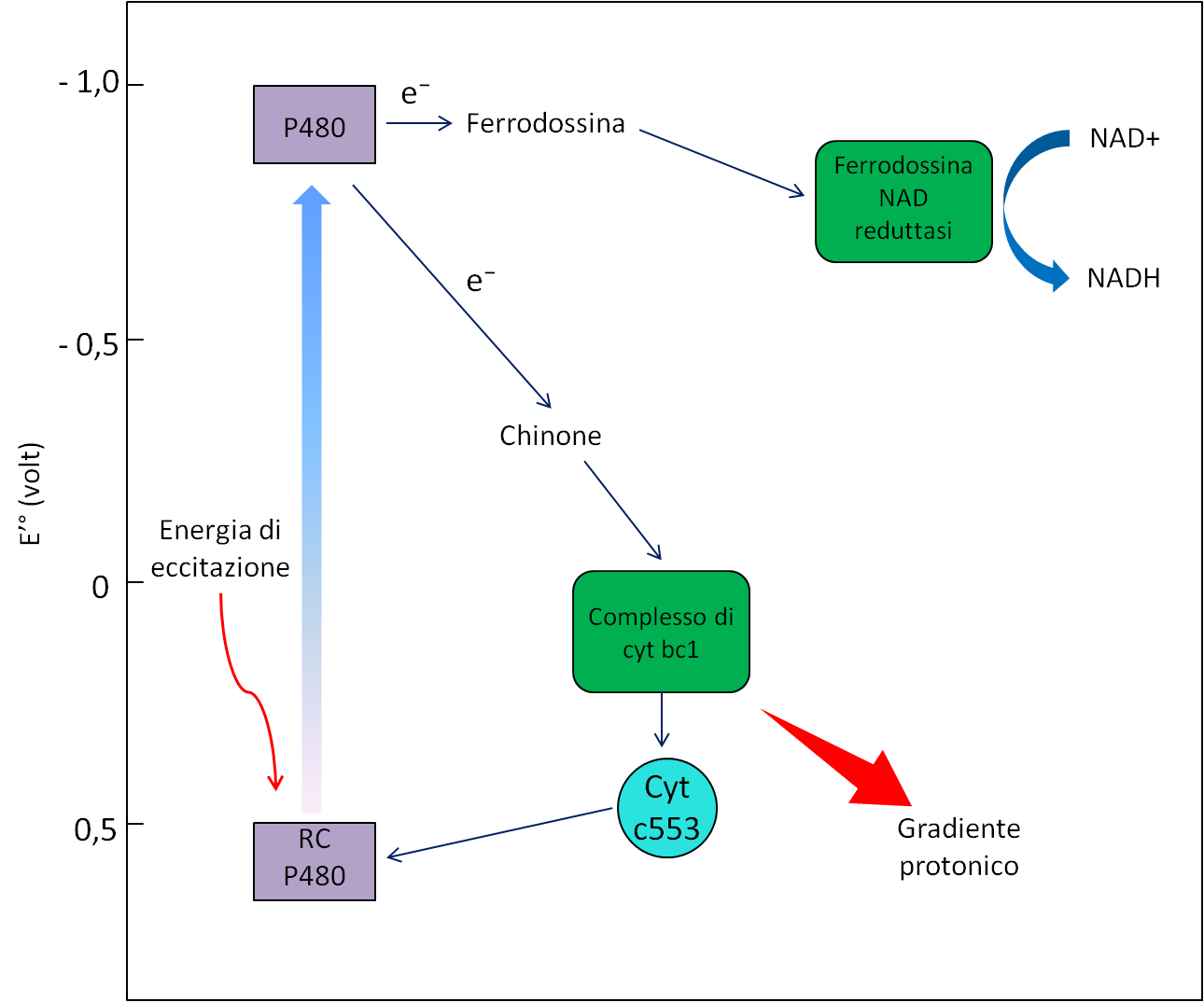
- Un percorso non ciclico verso la ferredossina e la ferredossina NAD reduttasi, con produzione di NADH (Fig. 13).
In entrambi i gruppi di batteri, il tragitto ciclico degli elettroni comporta il pompaggio dei protoni, che genera un gradiente elettrochimico rilevante per la sintesi di ATP.
Conclusione
Da ciò che è stato descritto in questo articolo, il meccanismo fotosintetico delle piante è importante per la loro sopravvivenza, ma soprattutto ricopre un ruolo decisamente essenziale per la vita in generale sul pianeta Terra, dal momento che, mediante la fotosintesi, le piante secernono ossigeno. Se consideriamo l’altro aspetto della fotosintesi, ossia l’assorbimento dell’anidride carbonica, non possiamo non affermare che la presenza delle piante gioca un ruolo fondamentale nel contrastare l’accumulo di CO₂ dovuto all’attività umana, responsabile dell’aumento delle temperature (quindi dell’effetto serra).
Per tale ragione le strategie migliori per salvare il pianeta dai cambiamenti climatici sono limitare l’abbattimento degli alberi, effettuare la riforestazione nelle zone rese desertiche dalla deforestazione eccessiva e, all’interno delle città, conservare i parchi e creare dei piccoli giardini sulle cime dei palazzi (Fig. 14). A proposito della realizzazione di tetti allestiti con alberi e piante, giardini pensili e terrazzi ricoperti di prati, c’è da dire che attualmente si sta diffondendo sempre di più nei centri urbani del mondo; New York, per esempio, ha deliberato l’obbligo dei “tetti verdi” per ogni edificio nuovo o in ristrutturazione. In Italia, Milano ha accettato il Piano di Governo del Territorio per il 2030 che, inoltre, offrirà la possibilità di rivalutare i tetti degli edifici come zone su cui mirare per uno sviluppo sostenibile della città.

Fonti
- David L. Nelson e Micheal M. Cox. 2014. “Introduzione alla biochimica di Lehninger”, Zanichelli
- Smith, A. L. 1997. “Oxford dictionary of biochemistry and molecular biology”, Oxford University Press
- Jeff Hardin, Gregory Paul Bertoni e Lewis J. Kleinsmith. 2014. “Becker il mondo della biologia”, Pearson
- R. B. Woodward, W. A. Ayer, J. M. Beaton, F. Bickelhaupt, R. Bonnett, P. Buchschacher, G. L. Closs, H. Dutler, J. Hannah, F. P. Hauck, S. Itô, A. Langemann, E. Le Goff, W. Leimgruber, W. Lwowski, J. Sauer, Z. Valenta, and H. Volz. 1960. ”The total synthesis of chlorophyll”, Journal of the American Chemical Society
- Ian Fleming. 1967. ”Absolute Configuration and the Structure of Chlorophyll”, Nature
- Robert Burns Woodward, William A. Ayer, John M. Beaton, Friedrich Bickelhaupt, Raymond Bonnett, Paul Buchschacher, Gerhard L. Closs, Hans Dutler, John Hannah, Fred P. Hauck, et al. 1990. “The total synthesis of chlorophyll a” Tetrahedron
- https://www.idroponica.it/fotosintesi-clorofilliana-piante_28-168.html
- https://e-l.unifi.it/pluginfile.php/54821/mod_resource/content/1/Lezione%208%20La%20fotosintesi%20%5Bmodalit%C3%A0%20compatibilit%C3%A0%5D.pdf
- http://www.benesserelongevitasalute.it/2017/03/30/alimenti-ricchi-di-clorofilla/
- http://www.ianua.com/patrizia/scuola/file/biologia/fotosintesi_clorofilliana_5.pdf
- Arms K. & Camp P. 1998. “Biologia”, Piccin
- Leonardi S. 2004. “Appunti di Ecologia Vegetale”, Dipartimento di Scienze Ambientali Università di Parma, Parma
- https://slideplayer.it/slide/571316/
- https://www.archiportale.com/news/2020/05/case-interni/milano-tetti-giardino-contro-l-inquinamento_76257_53.html
- https://www.cosedicasa.com/ristrutturare/giardino-sul-tetto-vantaggi-piante-prato-copertura-93788
- https://viaggionellascienza.it/materia/biologia/la-fotosintesi-clorofilliana/
1 commento su “Fotosintesi”
I commenti sono chiusi.