Il ruolo del piruvato nella fermentazione batterica
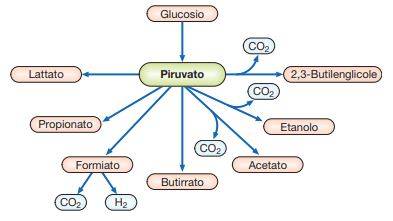
I batteri sono in grado di utilizzare numerose sostanze organiche, soprattutto carboidrati, tra le quali la più importante è sicuramente il glucosio. Nella fermentazione, il prodotto terminale dell’ossidazione del glucosio è rappresentato dall’acido piruvico (piruvato), una molecola alla quale vengono convertiti tutti i composti da 4-6 atomi di carbonio. L’ossidazione ulteriore del piruvato rappresenta una caratteristica biochimica che distingue i batteri tra di loro.
In ogni caso il piruvato, o un suo metabolita, funziona da accettore finale di idrogenioni che si accumulano in forma ridotta nel citoplasma batterico. La conversione del glucosio a piruvato avviene attraverso la via di Embden-Meyerhof e Parnas (EMP) e inizia con la fosforilazione del glucosio a glucosio-6-fosfato con il consumo di una molecola di ATP. Successivamente avviene l’isomerizzazione (fruttosio-6-fosfato) e una seconda fosforilazione (con il consumo di una seconda molecola di ATP), che porta alla formazione di fruttosio-1,6-difosfato. Questa molecola viene quindi scissa in due molecole di fosfogliceraldeide (molecola a 3 atomi di carbonio).

La deidrogenazione delle molecole di fosfogliceraldeide avviene grazie alla riduzione del NAD a NADH. L’energia liberata in questa fase viene utilizzata per fosforilare la fosfogliceraldeide ridotta generando, in questo modo, 2 molecole di acido 1,3-difosfoglicerico. A questo punto avviene la prima fosforilazione a livello del substrato in cui le due molecole di acido 1,3-difosfoglicerico cedono il loro gruppo fosfato all’ADP fosforilandolo ad ATP.
Il prodotto della reazione di idrolisi dell’acido 1,3-difosfoglicerico (acido 3-fosfoglicerico) viene isomerizzato ad acido 2-fosfoglicerico e dopo la perdita di una molecola di H2O si arriva alla formazione di 2 molecole di acido 2-fosfoenolpiruvico (PEP). In questa fase avviene una seconda fosforilazione a livello del substrato in quanto entrambe le molecole di acido 2-fosfoenolpiruvico cedono il loro radicale fosfato a due molecole di ATP. Le due fosforilazioni a livello del substrato che avvengono durante la fermentazione del glucosio a piruvato producono un totale di 4 molecole di ATP per molecola di glucosio ossidata, ma due molecole di ATP vengono consumate per trasformare il glucosio in fruttosio-1,6-difosfato, pertanto la resa netta della via EMP è di due molecole di ATP.
Il NADH, prodotto nella prima fase metabolica, rappresenta una molecola che deve necessariamente essere ossidata a NAD affinché la fermentazione possa avvenire in continuo. Esistono molti meccanismi di ossidazione del NADH a spese del piruvato e spesso i prodotti di ossidazione di queste reazioni caratterizzano in modo così specifico i batteri da essere utilizzate negli odierni sistemi di identificazione per distinguere tra loro le varie specie batteriche. In particolare, il piruvato può essere decarbossilato ad acetaldeide, che viene successivamente utilizzata per ossidare il NADH (fermentazione alcolica), oppure il piruvato può essere semplicemente ridotto ad acido lattico in modo da ossidare il NADH (fermentazione lattica).
Fermentazione lattica
La fermentazione lattica può essere omolattica, ovvero tutto il piruvato viene ossidato ad acido lattico (Streptococcus, Lactobacillus, ecc.), oppure eterolattica, dove il prodotto finale della fermentazione è rappresentato da una miscela complessa di acido lattico, etanolo e CO2 (batteri lattici, Leuconostoc). La fermentazione formica è tipica degli enterobatteri (Escherichia, Salmonella, Proteus ecc.).
Esistono due tipi di fermentazione formica, la fermentazione acido-mista e la butandiolica. I prodotti finali della fermentazione acido-mista sono rappresentati da una complessa miscela di acido lattico, acido acetico, acido succinico e acido formico. In presenza della formico-deidrogenasi, l’acido formico può essere ulteriormente trasformato in H2 e CO2. Nella fermentazione butandiolica, tipica dei generi Klebsiella, Enterobacter, Serratia e alcune specie di Bacillus, il butandiolo si forma per condensazione di due molecole di acido piruvico.
È importante sottolineare che nella fermentazione formica è possibile fosforilare una molecola di ATP durante l’ossidazione del NADH grazie alla presenza dell’acetil-CoA. L’acetil-CoA viene utilizzato per produrre acetil-fosfato, che successivamente dona il gruppo fosfato all’ADP. Nella fermentazione butirrico-butilenica, caratteristica del genere Clostridium (anaerobio stretto), dall’ossidazione del piruvato si ottengono H2O, CO2 e acetil-CoA. Quest’ultimo viene ridotto con produzione di acetone, isopropanolo e butanolo. Nella fermentazione propionica, presente nel genere Propionebacterium, il piruvato viene ossidato, attraverso una serie di reazione biochimiche, ad acido succinico. Questo, interagendo con l’acetil-CoA, produce una molecola di ATP, acido propionico e CO2.
Fonti
- Berg, J.M., Tymoczko, J.L., Stryer, L., Biochimica, 7a ed., Zanichelli, Bologna, 2012.
- Downs, D.M. (2006), «Understanding microbial metabolism», Annual Review of Microbiology, 60, p. 533.
- Hurst, C.J. et al. (a cura di), Manual of Environmental Microbiology, 2a ed., ASM Press, 2002.
- Nelson, D.L., Cox, M., I principi di biochimica di Lehninger, 5a ed., Zanichelli, Bologna, 2012.
- Russel, J.B., Cook, G.M. (1995), «Energetics of bacterial growth: balance of anabolic and catabolic reactions», Microbiological Review, 59, p. 285