I riarrangiamenti cromosomici sono anomalie strutturali che occorrono nelle sequenze di DNA cromosomali. Tali variazioni hanno svolto un ruolo importante nell’evoluzione dei genomi di molteplici forme di vita, tra cui l’essere umano, ma sono anche la causa di alcune malattie genetiche.
Anomalie cromosomiche
Le anomalie cromosomiche si dividono in due tipologie:
- Anomalie numeriche: sono presenti uno o più cromosomi in eccesso, oppure uno o più cromosomi sono assenti. Alcuni esempi sono la sindrome di Down o trisomia 21, la trisomia 18, la trisomia 13 e la sindrome di Turner;
- Anomalie strutturali: consistono in alterazioni della struttura dei cromosomi. Le anomalie strutturali sono sbilanciate se il cromosoma nativo è soggetto a delezioni, duplicazioni, inserzioni o sostituzione di sequenze di DNA. Le anomalie strutturali bilanciate corrispondono all’inversione o alla traslocazione di regioni cromosomali.
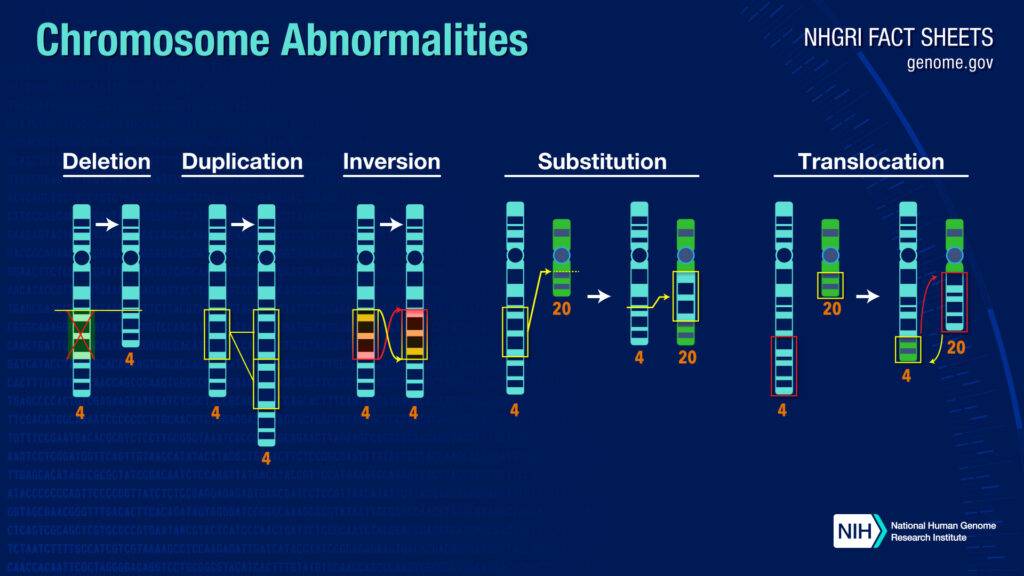
Delezioni e duplicazioni
Le delezioni cromosomali possono essere terminali oppure interstiziali. Nel primo caso, le estremità del cromosoma vengono accorciate. Nel secondo caso, avviene l’eliminazione di una regione presente all’interno del cromosoma.
Le duplicazioni cromosomali si distinguono principalmente in duplicazioni in tandem e inverse. Le duplicazioni cromosomali in tandem mantengono lo stesso verso di entrambe le sequenze identiche di DNA, mentre nel caso delle duplicazioni inverse un segmento cromosomale si orienta con verso opposto rispetto all’altro segmento.
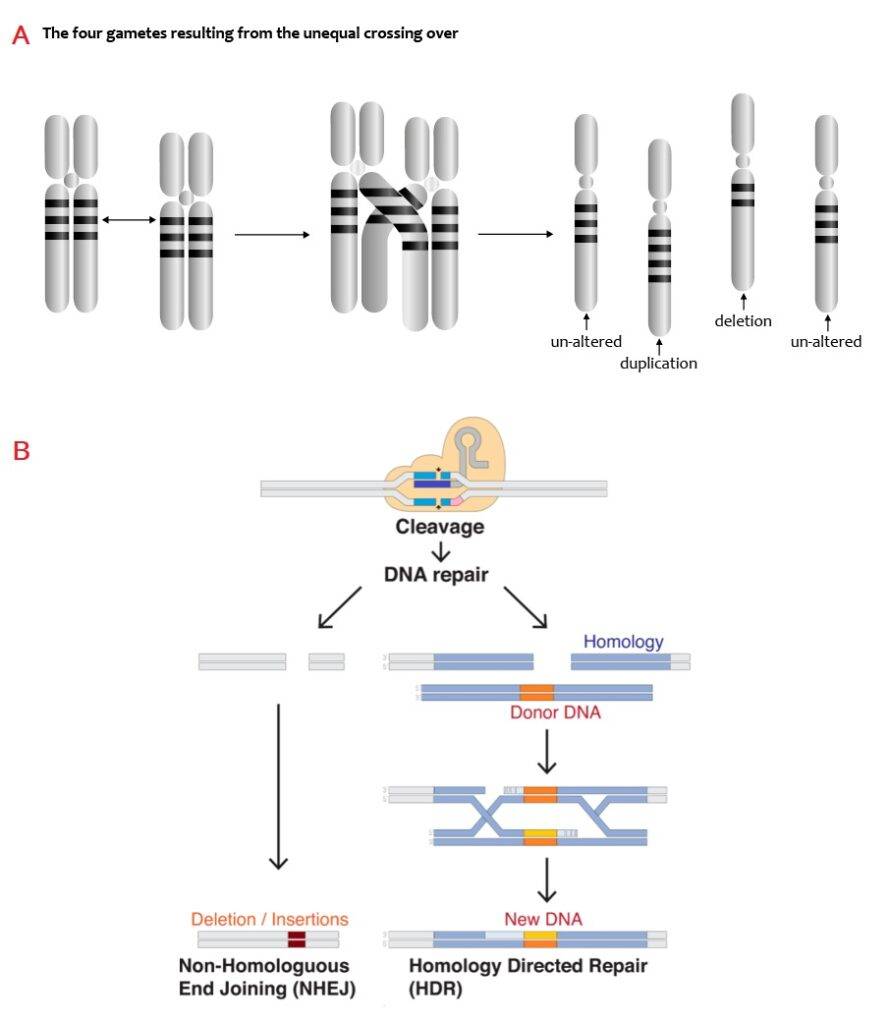
Delezioni e duplicazioni possono originarsi durante la meiosi o come conseguenza di rotture a doppio filamento del DNA (DNA Double-Strand Breaks – DSBs).
Nel primo caso, la ricombinazione omologa non allelica (Non-Allelic Homologous Recombination – NAHR) dà origine a duplicazioni o delezioni cromosomali in seguito al crossing-over.
Nel secondo caso, potenziali errori nel processo di giunzione delle estremità non omologhe (Non-Homologous End-Joining – NHEJ), causano inserzioni o delezioni di brevi sequenze di DNA. Il meccanismo NHEJ è distinto dalla riparazione diretta dall’omologia (Homology Directed Repair – HDR), per cui la cellula utilizza le sequenze di DNA presenti nel cromatidio fratello come “stampo” per ripristinare la porzione danneggiata di DNA.
Effetti delle delezioni
Le conseguenze fenotipiche di una delezione dipendono da quali geni sono localizzati nella regione eliminata. Se la delezione include il centromero, il cromosoma non andrà incontro a segregazione durante la meiosi o la mitosi e molto probabilmente sarà eliminato.
Molte delezioni sono letali in omozigosi perché tutte le copie di un gene contenute nei tratti di DNA rimossi sono assenti.
L’eterozigosi per una delezione può causare:
- aploinsufficienza: una sola copia di un gene non è sufficiente per produrre il fenotipo wild-type;
- pseudodominanza: l’unica copia del gene presente contiene una mutazione recessiva, che in assenza dell’allele wild-type si esprime a livello fenotipico.
Nell’uomo, alcuni esempi di malattie genetiche causate da delezioni sono la sindrome cri-du-chat, la sindrome Wolf-Hirschhorn e la sindrome Williams-Beuren.
Effetti delle duplicazioni
La presenza di copie aggiuntive di un gene impatta sul dosaggio genico, in quanto la quantità di proteine tradotte nella cellula è spesso direttamente proporzionale al numero di copie di geni.
Nell’uomo, alcuni esempi di malattie genetiche causate da duplicazioni sono la duplicazione parziale del braccio corto del cromosoma 4 (duplicazione 4p), la duplicazione parziale del braccio lungo del cromosoma 7 (sindrome da duplicazione 7q11.23) e la duplicazione parziale del braccio corto del cromosoma 9 (duplicazione 9p).
Le duplicazioni hanno svolto un ruolo importante nell’evoluzione degli eucarioti. Le copie esistenti di un gene sono essenziali per lo sviluppo e la fisiologia di un organismo. Le copie duplicate di un gene hanno maggiore possibilità di subire diverse mutazioni tra generazioni successive e, con la selezione naturale, di assumere delle nuove funzioni benefiche per l’organismo. Nell’uomo, i geni che codificano per le globine condividono lo stesso gene primordiale e hanno origine da duplicazioni sequenziali avvenute nel tempo.
Inversioni
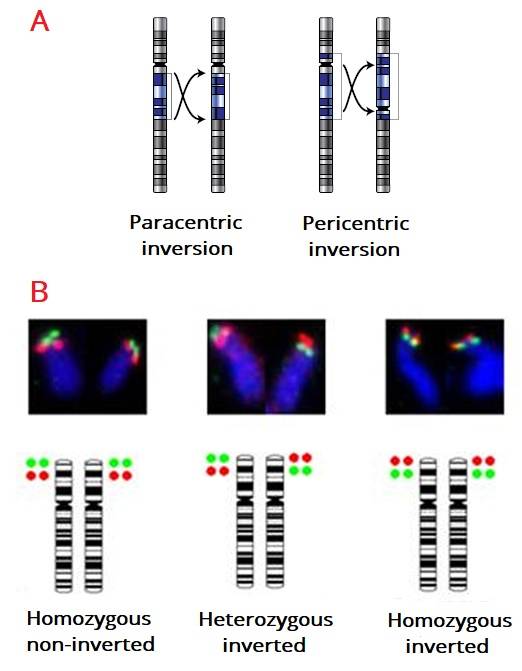
Le inversioni cromosomali sono paracentriche se non includono il centromero, oppure sono pericentriche se la porzione invertita del cromosoma comprende il centromero. Quando le interruzioni a doppio filamento del DNA sono localizzate sullo stesso cromosoma, se il DNA è riparato inserendo il segmento ruotato di 180°, si verifica un’inversione.
Effetti delle inversioni
A seconda della regione del DNA in cui si verifica, un’inversione potrebbe determinare:
- l’annullamento della funzione di un gene;
- un effetto posizione, se la regolazione di un gene dipende dalla regione cromosomale in cui è localizzato. Ad esempio, l’inversione di una regione del cromosoma X di Drosophila melanogaster determina lo spostamento del gene w+ in una regione cromosomale adiacente all’eterocromatina, con l’effetto che il gene viene silenziato.
Se un individuo è omozigote per un’inversione, con la meiosi i cromosomi omologhi si appaiano e si separano normalmente. Nel caso di un’inversione in eterozigosi, i cromosomi omologhi non si appaiano correttamente e si formano gameti ricombinanti, inadeguati nel dare origine alla progenie. Per questo motivo, l’inversione ha acquisito nel corso dell’evoluzione il ruolo di bloccare le ricombinazioni in alleli che si sono adattati ad un ambiente specifico.
Traslocazioni
La traslocazione è lo spostamento di sequenze di DNA tra cromosomi non omologhi, oppure all’interno dello stesso cromosoma. La traslocazione è diversa dal crossing-over, che consiste invece nello scambio di materiale genetico tra cromosomi omologhi. La traslocazione non reciproca o sostituzione è il trasferimento di DNA da un cromosoma ad un altro, senza uno scambio reciproco.
La traslocazione Robertsoniana è un tipo di traslocazione che coinvolge due cromosomi acrocentrici, in cui il centromero è molto vicino alle estremità del cromosoma. Lo scambio di DNA produce un cromosoma metacentrico e un cromosoma molto piccolo che viene eliminato.
Le rotture a doppio filamento del DNA sono il prerequisito per le traslocazioni.

Effetti delle traslocazioni
Le traslocazioni condizionano il fenotipo con modalità diverse:
- con l’effetto posizione, la regolazione dell’espressione di un gene trasferito in un cromosoma diverso può variare;
- la traslocazione di una sequenza di DNA all’interno di un gene potrebbe condurre al silenziamento dell’espressione genica.
Le delezioni accompagnano spesso le traslocazioni, come per la traslocazione Robertsoniana, che implica la riduzione del numero globale di cromosomi.
Gli effetti delle traslocazioni nella formazione dei gameti dipendono dal tipo di traslocazione. In linea generale, in un individuo eterozigote per una traslocazione reciproca, circa la metà dei gameti prodotti con la meiosi è funzionale, mentre l’altra metà non è vitale perché il corredo cromosomico è sbilanciato.
Fonti
- https://www.ncbi.nlm.nih.gov/books/NBK115545/
- https://www.genome.gov/about-genomics/fact-sheets/Chromosome-Abnormalities-Fact-Sheet
- http://opengenetics.net/Files/MRUOpenGeneticsLectures//Ch17-Chromosome_rearrangements.pdf
- R.J. McKinlay Gardner, D.J. Amor (2018), «Gardner and Sutherland’s – Chromosome Abnormalities and Genetic Counseling – 5th ed.», Oxford University Press, pp. 474-491
- B.A. Pierce (2020), «Genetics: A Conceptual Approach – 7th ed.», MacMillan Learning, chap. 8.
- Immagine in evidenza: https://www.flickr.com/photos/zeissmicro/14256070350
1 commento su “Riarrangiamenti cromosomici”
I commenti sono chiusi.