Caratteristiche
Il batterio Francisella tularensis è un coccobacillo Gram negativo pleomorfo (Fig. 1), aerobio obbligato, asporigeno, provvisto di capsula, immobile, ossidasi e catalasi positivo, responsabile di un’antropozoonosi conosciuta come tularemia.

Francisella tularensis presenta una capacità infettante assai elevata (da 1 a 10 batteri del tipo A bastano per provocare la malattia in svariate specie di animali da laboratorio), per tale ragione i laboratori che lavorano con campioni potenzialmente contaminati da tale microorganismo dovrebbero dotarsi di un livello di sicurezza 2, o addirittura 3, se manipolano grandi quantità di questo batterio.
Esistono due sottospecie di F. tularensis, ovvero Francisella tularensis tularensis (tipo A) e Francisella tularensis holarctica (tipo B); il primo, che è il più infettivo, si trova principalmente in America Settentrionale, ed è letale sia per l’essere umano che per il topo; il secondo è diffuso soprattutto nei roditori dell’Europa, dell’Asia e del Nord America, e causa una malattia più lieve.
I batteri del genere Francisella sono resistenti e capaci di persistere nel fango, nell’acqua e nei carcami di animali in decomposizione. Francisella tularensis riesce a sopravvivere e a moltiplicarsi all’interno delle amebe allo stesso modo delle specie di Legionella spp. Tale proprietà è vantaggiosa per la sua sopravvivenza e replicazione in ecosistemi ambientali come acqua e melma.
Il primo isolamento di questo microorganismo risale al 1912 ad opera di George McCoy, il quale stava investigando su una patologia simile alla peste bubbonica che colpiva gli scoiattoli; egli chiamò il batterio Bacterium tularense, dal nome della contea di Tulare, in California, luogo in cui era stato osservato il focolaio per la prima volta. Nel 1919, il batteriologo statunitense Edward Francis determinò che l’infezione chiamata “febbre del cervo volante” era la stessa malattia, e la nominò “tularemia”. Proprio in onore di questo scienziato, nel 1947 il batterio è stato designato come un nuovo genere e definito Francisella; precedentemente era stato collocato nel genere Pasteurella e in seguito nel genere Brucella. Francis comprovò che l’infezione riguardava i conigli selvatici e incautamente scoprì che era altamente infettiva, dal momento che sia lui che tutti i suoi assistenti del laboratorio ne rimasero contagiati. Questa caratteristica spinse a studiare il suo potenziale come arma biologica, ricorrendo alla sperimentazione umana non volontaria da parte del Giappone tra civili, politici, prigionieri militari, e il suo possibile uso nei conflitti durante la Seconda Guerra Mondiale. Tra il 1950 e il 1960, negli Stati Uniti iniziò la sperimentazione umana volontaria con soldati civili e detenuti. Si presume che gli scienziati dell’Unione Sovietica svilupparono un ceppo resistente al vaccino che testarono come arma biologica nel 1982-1983.
Filogenesi
Dominio Prokaryota
Regno Bacteria
Phylum Proteobacteria
Ordine Thiotrichales
Famiglia Francisellaceae
Genere Francisella
Specie F. tularensis
Epidemiologia
Francisella tularensis è ubiquitaria alla latitudine tra 30° e 70° nell’Emisfero Boreale, a partire da alcuni territori a nord del Circolo Polare Artico fino ai paesi come la Scandinavia, l’America del nord, la Russia e il Giappone (Fig. 2).

L’infezione è conosciuta sia nei vertebrati che negli invertebrati (almeno 145 e 111 specie rispettivamente). Nel primo gruppo sono inclusi soprattutto i mammiferi, tra cui lagomorfi (conigli e lepri), roditori, mustelidi, insettivori, carnivori, ungulati e marsupiali; a questi si aggiungono ulteriori classi di vertebrati, ossia uccelli, rettili, anfibi e pesci. Nel secondo gruppo rientrano le zecche, i tafani, le zanzare e le pulci.
Nei periodi in cui la malattia non è epidemica, gli organismi che costituiscono i serbatoi dell’infezione sono le zecche appartenenti al genere Ixodes, che trasmettono il batterio in maniera trans-stadiale, cioè dallo stadio di larva a ninfa e da ninfa ad adulto. Anche alcune specie di lagomorfi e roditori possono diventare serbatoi del microrganismo in quanto sono poco suscettibili all’infezione, tra questi abbiamo i conigli del genere Sylvilagus, i castori e i topi muschiati del Nord America e i topi campagnoli europei. Al contrario, le lepri europee non raffigurano un serbatoio dal momento che manifestano una forma acuta setticemica con elevata mortalità. Il coniglio europeo selvatico, invece, è parzialmente resistente all’infezione.
Morfologia delle colonie
Considerata l’esigenza di cisteina o di composti sulfidrilici (tioli) in quantità apprezzabile, i terreni più adatti per la coltura della Francisella tularensis sono l’agar sangue con aggiunta di glucosio e cisteina e il Thayer-Martin (una variante di agar cioccolato addizionato con vancomicina, colistina e trimethoprim), incubati a 35°C in atmosfera contenente 10% di anidride carbonica (CO₂). Nel giro di due o quattro giorni si formano delle colonie bianco-grigiastre (Fig. 3), viscose se il batterio è in fase S, oppure rugose se è in fase R. Su agar sangue si può notare un sottile alone verde di emolisi che circonda le colonie.

Fattori di virulenza
F. tularensis possiede l’abilità di penetrare e replicarsi nei macrofagi e ciò raffigura la maggiore proprietà patogenetica del batterio. I geni che permettono la sopravvivenza e la divisione del microorganismo all’interno dei macrofagi sono stati identificati impiegando la mutagenesi basata sui trasposoni. Tra questi abbiamo il fattore di trascrizione MglAB, fondamentale per la crescita nei macrofagi della F. novicida ed è espresso anche dalla F. tularensis; tale molecola regola l’espressione di svariati geni necessari per la replicazione nei macrofagi e per la virulenza, inclusi i geni per la crescita intracellulare (iglA, iglC e iglD ) e i geni determinanti la patogenicità (pdpD e pdpA). Questi loci sono localizzati in un tratto del genoma di circa 30 kb che esibisce le caratteristiche di un’isola di patogenicità, tra cui il basso contenuto di guanina e citosina e la presenza di geni di virulenza. Questa regione è stata designata l’isola di patogenicità della Francisella (FPI).
MglAB presenta un’elevata omologia con una proteina di Escherichia Coli, ovvero SspAB (un regolatore della risposta alla mancanza urgente dei nutrienti), e tra l’altro si ipotizza possa controllare l’espressione di una fosfatasi appartenente al gruppo di proteine in cui è compresa AcpA. Questa è una fosfatasi acida che inibisce lo scoppio respiratorio (cioè il processo in cui alcune cellule liberano gli intermedi reattivi dell’ossigeno) e mostra un’attività a fosfolipasi C. In esperimenti con ceppi di F. novicida mutanti per AcpA si è scoperto che l’assenza di questa proteina rende il batterio suscettibile all’uccisione intracellulare, in quanto l’evasione dal fagosoma è ritardata. Questa molecola, inoltre, sembra giocare un ruolo cruciale nella sopravvivenza della Francisella tularensis nel tratto respiratorio. Un ulteriore fattore implicato nella sopravvivenza nei macrofagi è la proteina minD, avente due probabili ruoli: operare come una pompa per i radicali liberi e compiere una funzione durante la replicazione batterica nelle cellule.
Trasmissione e patogenesi
Come abbiamo già descritto all’inizio, Francisella tularensis è l’agente eziologico della tularemia, una malattia che colpisce sia animali che persone. Conigli, lepri e roditori sono particolarmente suscettibili e spesso muoiono in gran numero durante le epidemie. Nelle persone la patologia è legata agli ambienti rurali, di conseguenza riguarda soggetti che trascorrono la maggior parte del loro tempo nella campagna, soprattutto cacciatori ed escursionisti. La trasmissione del batterio si può verificare in molteplici modalità:
- Contatto diretto con animali infetti;
- Manipolazione delle carcasse di animali infetti;
- Ingestione di acqua contaminata;
- Inalazione di polveri provenienti da terreno, fieno e grano contaminati;
- Puntura di tabanidi, culicidi e varie specie di zecche, tra cui Amblyomma americanum e Dermacentor variabilis (Fig. 4);
- Mangiando carni contaminate poco cotte;
- Attraverso il morso di alcuni animali (coyote, scoiattolo, puzzola, maiale, cinghiale, gatto e
cane), ma è un evento meno frequente.
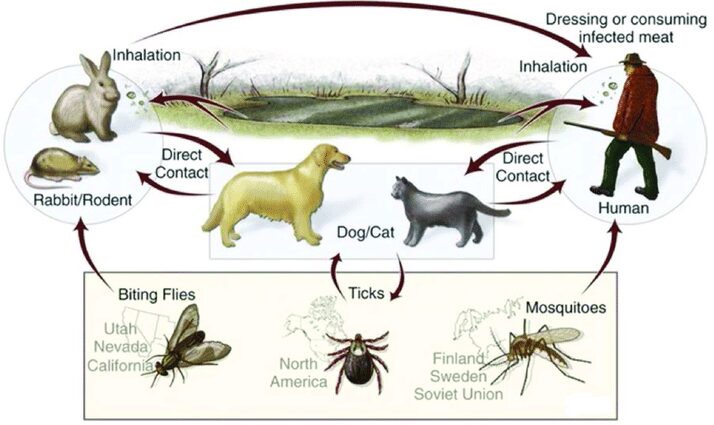
La figura 5 ci fornisce una raffigurazione del ciclo della F. tularensis dove sono indicate le nazioni e i continenti associati alla trasmissione del batterio da parte di uno specifico vettore.

Per quanto concerne il quadro clinico della tularemia, negli animali e negli esseri umani si osservano sintomi differenti. Iniziando dagli animali, nel gatto, che è l’animale domestico più sensibile, le manifestazioni sono depressione spiccata, linfadenite regionale (a livello dei linfonodi faringei, cervicali o mesenterici) o generalizzata, splenomegalia, epatomegalia, ittero e panleucopenia. A volte avviene la formazione di ulcere superficiali nella bocca e sulla lingua.
Nel cane, che di solito è resistente alla malattia, i sintomi sono generalmente anoressia, letargia, diarrea, brividi, mialgia e ipertermia. In alcuni casi può presentare tonsillite, ascessi sottocutanei, ingrossamento dei linfonodi superficiali, uveite e congiuntivite.
Nella pecora la patologia è caratterizzata da depressione, ipertermia, rigidità della camminata, diarrea e mortalità. I primi due sintomi si manifestano anche nel cavallo, insieme a mancata coordinazione e dispnea.
Negli animali selvatici (come lepri, cani della prateria, coyoti e castori) la malattia può essere asintomatica fino alla morte, oppure si può avere la comparsa di anoressia, letargia, splenomegalia, debolezza muscolare e apatia, che sono seguiti dalla morte nel giro di due o tre giorni.
La tularemia nell’essere umano inizia con febbre, brividi, cefalea e mialgia, dopo un periodo di incubazione di tre o cinque giorni. In base alla modalità di trasmissione, si riscontrano dei quadri clinici dissimili:
- Forma ulceroghiandolare, che raffigura la manifestazione più assidua, è distinta da una lesione cutanea dolorante e arrossata nel sito di inoculazione; questa si trasforma in una papula che si ulcera entro due giorni (Fig. 6A). Se è localizzata su arti inferiori, collo e viso, normalmente è secondaria alla puntura della zecca. Se invece si trova sugli arti superiori, nell’80% dei casi è legata alla manipolazione di carcasse. Questa lesione può essere preceduta da linfadenite satellite (Fig. 6B), in cui i linfonodi sono dolenti, con periadenite (infiammazione del tessuto connettivo molle che circonda le ghiandole linfatiche) e vanno incontro a colliquazione.
- Forma ghiandolare, che è simile a quella ulceroghiandolare ma senza la formazione dell’ulcera (Fig. 6C). Si acquisisce di solito attraverso il morso delle zecche o dei tabanidi, tramite il contatto con l’acqua contaminata oppure maneggiando animali morti o malati.
- Forma oculoghiandolare, caratterizzata da congiuntivite purulenta, dolorosa e unilaterale. La mucosa della congiuntiva appare iperemica, ricca di granuli giallastri e talvolta ulcerazioni (Fig. 6D). Si verifica in seguito all’entrata del batterio nell’occhio.
- Forma orofaringea, si manifesta con mal di gola, disfagia, ulcere orali, tonsillite e rigonfiamento dei linfonodi cervicali (Fig. 6E e 6F). È causata dall’ingestione di cibo e acqua contaminati.
- Forma setticemica, con febbre elevata e assenza di ulcerazioni e linfadenite. Colpisce specialmente il personale di laboratorio che ha effettuato riscontri autoptici su animali infetti.
- Polmonite, è la forma più grave. È rara in Europa ma diffusa negli Stati Uniti d’America, dove sono stati segnalati casi in individui che hanno respirato polveri contaminate durante la tosatura dei prati. I sintomi sono febbre alta, tosse secca, malessere, dolore toracico, dispnea ed eccezionalmente emoftoe. Può insorgere in seguito a diffusione ematogena e si appura nel 30%-80% delle forme setticemiche.
- Meningite, che è una manifestazione inconsueta nell’infezione da F. tularensis. Il liquido cerebrospinale appare ricco di cellule, in particolare linfociti, e si ha ipoglicorrachia e iperproteinorrachia, cioè riduzione del glucosio e aumento delle proteine rispettivamente.

Immunologia e anatomia patologica
Dopo tre o cinque giorni dall’inoculazione, il batterio prolifera a livello cutaneo, favorendo la creazione della papula che poi diventa ulcera, si diffonde ai linfonodi regionali e, per via linfatica ed ematica, può raggiungere gli altri organi. Microscopicamente le lesioni espongono una necrosi suppurativa focale che ricorda la reazione granulomatosa causata da Mycobacterium tuberculosis; l’area centrale di necrosi contiene soprattutto granulociti neutrofili e macrofagi. Possono essere coinvolti anche i fibroblasti, i quali circoscrivono la reazione infiammatoria, ed è possibile individuare cellule epiteliodi e cellule di Langhans (cellule giganti multinucleate di 40-50 μm di diametro i cui i nuclei formano un cerchio oppure un semicerchio a ferro di cavallo).
La persistenza del microorganismo nei macrofagi ha come conseguenza una risposta immuno-mediata e cellulo-mediata prolungata. Ciononostante, è difficile che i ceppi virulenti capsulati vengano lisati anche in presenza di anticorpi osponizzanti, dal momento che la capsula è ricca di lipidi. Dopo la replicazione nei macrofagi, Francisella tularensis induce la sintesi di radicali liberi, il rilascio di TNF-α e l’attivazione della caspasi 3 che promuove l’apoptosi della cellula ospite (la capacità di stimolare la morte cellulare programmata è una caratteristica comune ad altri batteri Gram negativi, come Salmonella, Shigella, Yersinia e Legionella). Queste reazioni hanno come risultato la genesi di necrosi e granulomi nei tessuti.
In uno studio del 2013, pubblicato su APMIS, i ricercatori eseguirono l’analisi istopatologica di 17 biopsie linfonodali, che rivelarono la comune morfologia di un infiltrato infiammatorio misto con necrosi caseosa, granulomi contenenti linfociti, istiociti epiteliodi e rare cellule giganti, ed infiammazione estesa oltre la capsula dei linfonodi (Fig. 7).

Metodi di identificazione
Poiché la tularemia è una malattia insolita e i sintomi possono essere scambiati per altre patologie più comuni, la diagnosi risulta complicata. Ragion per cui è importante condividere col proprio medico curante ogni eventuale esposizione, per esempio punture di zecche e di tabanidi, o contatti con animali malati o morti.
Ciò che può far sorgere il sospetto di tularemia sono la provenienza da una zona in cui la patologia è endemica e il dato epidemiologico (la manipolazione di carcasse di animali, l’ingestione di carne poco cotta, l’ingestione di acqua potenzialmente contaminata). Nel caso in cui un individuo presenta la forma ulceroghiandolare, bisogna eseguire la diagnosi differenziale con le adeniti da piogeni, la malattia del graffio del gatto (causata dal batterio Bartonella henselae), la febbre da morso di ratto (dovuta a due batteri Gram negativi presenti nell’orofarigne del ratto, ovvero Streptobacillus moniliformis e Spirillum minor) e l’infezione da Pasteurella. Per quanto riguarda le forme setticemiche, sono contraddistinte da un quadro clinico somigliante a quello di infezioni come salmonellosi, brucellosi, malaria, polmoniti batteriche e polmoniti atipiche. Le colture effettuate sul materiale estratto dalle lesioni cutanee o dai linfonodi di rado permettono di isolare il patogeno, in quanto i siti di infezioni si autosterilizzano prematuramente. Inoltre, nell’essere umano la batteriemia è alquanto infrequente, contrariamente agli animali.
È possibile ricercare F. tularensis nell’espettorato, nel tampone faringeo e congiuntivale coltivandola nell’apposito terreno. Ulteriori metodi per individuare il microorganismo sono la reazione a catena della polimerasi (PCR), il test sierologico e la microagglutinazione; quest’ultima funziona mettendo il siero del paziente a contatto con batteri uccisi all’interno di una soluzione alcolica. Questo test permette di evidenziare la presenza di anticorpi di classe IgM che riconoscono gli antigeni capsulari, che di solito compaiono dopo circa dieci giorni dall’inizio della malattia e possono mantenersi per anni. Se un soggetto ha un’infezione da Brucella, Yersinia o Proteus OX19, si possono verificare reazioni crociate, e quindi falsi positivi. Oltre alle metodiche elencate, esiste anche un saggio immunoenzimatico per ricercare anticorpi IgG, IgM e IgA, più sensibile rispetto al test di agglutinazione. La diagnosi sierologica definitiva è tale se si rimarca un numero quattro volte o più del titolo in due determinazioni attuate a distanza di 7-10 giorni.
Terapia e prevenzione
La terapia della tularemia si fonda sull’impiego degli amminoglicosidi, tra cui streptomicina e gentamicina, che raffigurano gli antibiotici di prima scelta data la loro attività battericida verso F. tularensis. Nel caso in cui insorge la meningite, è necessaria la somministrazione di cloramfenicolo o doxiciclina insieme alla streptomicina, dal momento che quest’ultima difficilmente riesce a superare la barriera emato-encefalica.
È possibile utilizzare anche i fluorochinoloni come telitromicina e ciprofloxacina, anch’essi dotati di attività battericida. Oltre agli antibiotici, la cura della tularemia richiede anche il drenaggio dei linfonodi colliquati.
Normalmente il trattamento dura dai 10 ai 21 giorni a seconda dello stadio della malattia e del trattamento applicato.
Per quanto concerne la profilassi, nelle aree endemiche è opportuno l’uso di guanti protettivi quando si sta in contatto o si maneggiano animali malati, moribondi o morti, applicare repellenti contro gli insetti vettori, evitare di falciare l’erba dove sono presenti animali morti, cuocere bene la selvaggina. Nel controllo delle epidemie legate all’acqua, si attesta l’efficacia della clorazione. Per il personale di laboratorio che maneggia colture di F. tularensis è raccomandata la vaccinazione basata su microorganismi vivi attenuati, che offre un’adeguata immunità.
Poiché la tularemia non si trasmette da individuo a individuo, non occorre mettere in isolamento i pazienti ricoverati.
Arma biologica
Ci sono diversi fattori che sostengono l’impiego di Francisella tularensis come arma biologica, ovvero la bassa concentrazione sufficiente a provocare l’infezione, la semplicità di propagazione (per via aerea e tramite l’acqua) e il potere patogeno del batterio.
Date le sue proprietà, la F. tularensis è stata reputata come arma biologica a partire dal 1932, perciò come potenziale rischio sia dagli Stati Uniti d’America che dalla ex Unione Sovietica. Negli USA il Center Disease of Control (CDC) ha inserito il microorganismo nella classe A degli agenti biologici insieme al Poxvirus del vaiolo, a Bacillus anthracis, a Yersinia pestis, a Clostridium botulinum e ai virus delle febbri emorragiche (Ebola, Marburg, Lassa, Junin), dal momento che sono agenti che si spargono agevolmente, possono determinare alta mortalità, suscitare terrore e “scompiglio sociale”.
Si suppone che le epidemie di tularemia che colpirono migliaia di soldati tedeschi e sovietici durante la seconda guerra mondiale fossero connesse alla diffusione dolosa del batterio. Con le tecniche di ingegneria genetica si è cercato di creare ceppi di F. tularensis inalterabili dal cloramfenicolo e dalle tetracicline. Stipiti virulenti resistenti alla streptomicina sono stati saggiati durante alcuni studi il cui scopo era il bioterrorismo negli Stati Uniti e nell’Unione Sovietica.
Fonti
- Fabbi M., Magnino S., Marone P. 2005. “Trattato sulle infezioni e tossinfezioni alimentari”, Selecta Medica
- Gökcen Dinc, Hayati Demiraslan, Mehmet Doganay. 2017. “Unexpected Risks for Campers and Hikers: Tick-Borne Infections”, Int J Travel Med Glob Health
- Encyclopædia Britannica, IncJ. V. Hirschmann. 2018. “From Squirrels to Biological Weapons: The Early History of Tularemia”, Am J Med Sc
- Nrusingh P. Mohapatra, Ashwin Balagopal, Shilpa Soni, Larry S. Schlesinger, and John S. Gunn.. 2007. “AcpA Is a Francisella Acid Phosphatase That Affects Intramacrophage Survival and Virulence”, Infect Immun
- Anna Brotcke, David S. Weiss, Charles C. Kim, Patrick Chain, Stephanie Malfatti, Emilio Garcia, Denise M. Monack. 2006. “Identification of MglA-Regulated Genes Reveals Novel Virulence Factors in Francisella tularensis”, American Society for Microbiology
- Amina Nemmour, Adzreil Bakri, Claude A Fischer and Yves Brand. 2019. “Paediatric oropharyngeal tularaemia requiring surgical intervention”, BMJ Case Reports
- Ersin Tuncer, Binnur Onal, Gulcin Simsek, Sahande Elagoz, Ahmet Sahpaz, Selcuk Kilic, Emine Elif Altuntas and Aysegul Ulu Kilic.2013.“Tularemia: Potential role of cytopathology in differential diagnosis of cervical lymphadenitis: Multicenter experience in 53 cases and literature review”, APMIS